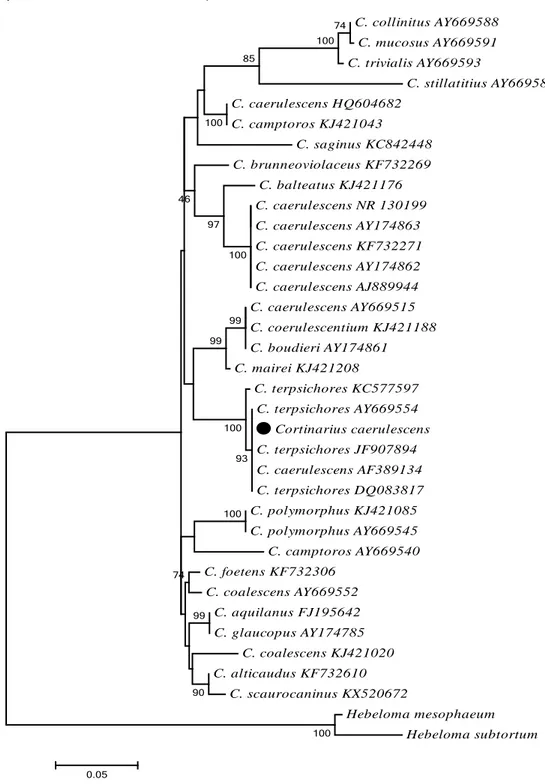
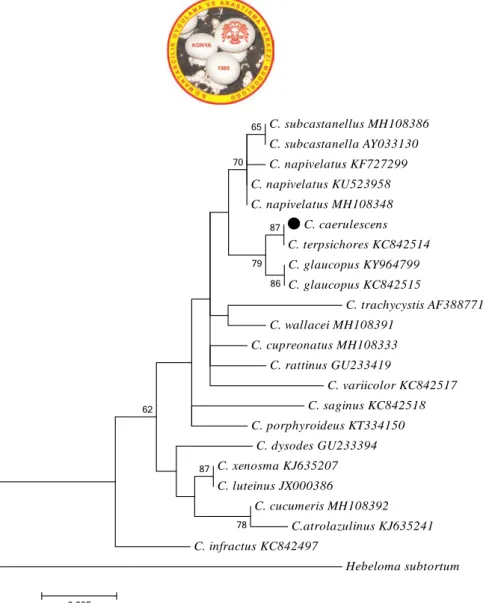
Phylogenetic and Taxonomic Studies on Cortinarius caerulescens (Schaeff.) Fr. a New Record for Turkish Mycota
Tam metin
Şekil

Benzer Belgeler
At day 3 of nicotine administration, an 82% increase in the expression of the longer form of Homer2a ⁄ b was observed by real-time RT-PCR analysis (P < 0.05; Fig. 1A) within
Bu yazida dieffenbachia bitkisinden yarim yaprak yedikten sonra ağizda yanma ve şişlik gelişen üç yaşindaki çocuk hasta sunularak ev ortaminda sik kullanilan süs
黃帝外經 補瀉陰陽篇第七十三 原文
The power capacity of the hybrid diesel-solar PV microgrid will suffice the power demand of Tablas Island until 2021only based on forecast data considering the
Çalışmanın ikinci kısmında gayesi insan davranışlarını açıklama ve anlama olan modern psikoloji biliminin verileri ışığında bu bilgiler ele alınıp analiz edilecek,
- Okul bazında, eğitim sürecine katılan tüm personelin BT konusunda eği timi ve onların bu konudaki gelişimleri ile ilgili faaliyetlerin planlanması, ilgili
Ding, Linear complexity of generalized cyclotomic binary sequences of order 2, Finite Fields Appl. Ding, Autocorrelation values of generalized cyclotomic sequences of order two,
In this study, for predicting Level 1 subfamilies of GPCRs, a novel method for obtaining class specific features, based on the existence of activating ligand specific patterns,
