Comparison of the Anti-inflammatory Effects of Proanthocyanidin, Quercetin, and Damnacanthal on Benzo(a)pyrene Exposed A549 Alveolar Cell Line
Tam metin
Şekil


Benzer Belgeler
In the current study, treatment with pravastatin inhibited markedly the L-NAME- induced high total and LDL cholesterol levels although statins are demonstrated to have no lipid
In conclusion, pneumoperitoneum performed after thoracotomy does not increase the inflammatory reaction and oxidative stress caused by thoracotomy and is an effective
Results: It was determined that HP extract treatment increased the expression of apoptotic proteins (AIF, GADD153, GRP78, caspase-3, Bax) and cell cycle G2 / M checkpoint
No significant difference was found between lesio- nal tissues and control tissues in terms of TNF- α and IFN-γ but Fas R was statistically significantly higher in the patient
*p<0.05 significant compared to control and # : p<0.05 significant compared to standard, MELR: Methanolic extract of Limnophila repens, SEM: Standard error of the mean..
Cytotoxic activity of plant extract on different cancer cell lines The cytotoxicity of the crude ethanolic extract from the flowering parts of C.. solstitialis at
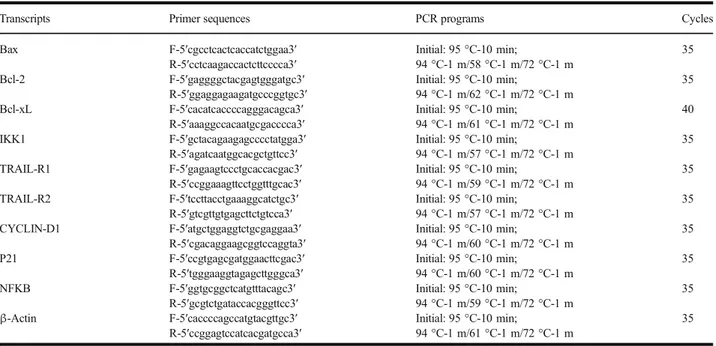
In this study, we measured the expression levels of P53, BAX, BCL-2, CAS3, CAS9, BIRC, and PPIA (housekeeping) genes related to apoptosis on A549 human lung carcinoma
Similarly, in our study, TNF-α and IL-6 levels were found to be higher in patients with influenza than in the control group and were even higher in fatal patients1. IL-1β levels
