iDRARDA BULUNAN PÜRiN DERiVATLARININ MiKROBiYEL PROTEiN
KATKısı VE YEM TÜKETiMiNiN ÖLÇÜMÜNDE KULLANIMI
Şule Kaya'
The Use of Urinary Purine Dcrivatives to Estimate Microbial Protein Supply and Feed Inıake
Summary: The urinary excretion ol allanloin was lirsl suggested as an index ol rumen microbial protein yield about 30 years a9O. Major progress towards estabUshing a method to relate purine derivatives excretion to micrcbla! protein yield has only been made In recent years. So that. the blochemistry ol lhe purines. the factors eHecting the amount ol purine derivatives excreled in the urine and the diHerent approches to estlmate recovery ol Ihese denvatives ori ginating from differenl sources has been explained in order to gulde lor who is tnleresled in Ihis subject
Key Words :Purine derivatives, microblal protein. rumen
Özet: Yaklaşık 30 yıı önce, idrarla atılan allantoinin rumendeki mikrobiyel protein Oretlminin bir göstergesi olduğu ilk kez ileri sOr\ildO. Ancak pOrin derivaUannın atılımı ile mikrobiyel protein Orelimine Ilişkin bir melot ortaya koymaya yö nelik büyOk gelişmeler son yıllarda kaydedildi. Bu nedenle bu konuyla Ilgiıenenleri aydınlatmak amacıyla, pOrinlerın bi
yokimyası. idrarta atılan pOrin derivaılarının miktarını etkileyen faktörler, degişik kaynaklardan orijin alan bu derivaliarın
geri alınımını lespit elmek Için kullanılan larKlı ya!daşımlar açıklandı. Anahtar Kelimeler: POrin derivaUan, mikrobiyel proleln, rumen
Giriş
Hayvan ve bitki hücrelerinin temel yapılarından olan pürin ve pirimidlnıer değişik bileşimlerde bu lunurlar. ATP gibi bileşimieri çabuk değişime uğ rayabilirlerken, DNA gibi bileşimlerin kararlı olması dikkat çekicidir. Purin biyokimyasında purin bazını oluşturan en önemli yapı. ribeı yada deoksiriboı ve fosforlk asitten oluşan nukleoliddlr. Önemli pürin bazları adenin, guanin, hipoksantin ve ksanlln'dir. Bu yapıların ribeılannın, metabolizmada meydana geldiği bilinmektedir. Adenin ve guanin genelde memeliierin dokularında bulunmazlar. Fakat ser best ksantin ve hipoksantin purinlerin yıkımındaki ara maddelerdir. Ünk asit bir divalent asiilir fakat pH 7 civarında monobazik tuzlann oluştuğu ikinci ayrılma sabitesi çok küçOktür (Zöllner, 1982).
Nükleik asitlerin metabolizması, nükleolidlerin devamh olarak nükleozld ve serbest bazlara yı kımlandı{ıı substrat stklusunu içerir. Oluşan nük leozld ve bazlar yenı nukleolidlerirı sentezlenmesl için daha sonra da kullanıhrlar.
Gcliı Tıırihi: 30.12.1997 •
I.S.U. Vclcnncr Fakıllıesi. Fizyoloji Anabilim Dalı. KONYA.
Pürinlerin Hücre Içerisine Alınması
Hücrelerin ihtiyaçlarını karşıladıklan pürin kay· nakları arasında bir denge kurulmuştur. Bu kay nakların ilki, mikrobiyel veya gıdal kökenli ekzojen olarak vücuda ahnan purinler, diğeri ise yenıden sentezlenen pOrinlerdlr.
parinler yeniden sentezlenirken ; amonyak, as partik asit ve glisin gibi çok basit ön maddeleri kul lanırlar ve bu dokuların pürinler için spesifik ih· tiyaçları olmadl{ıt anlamına gelirse de, bu işlem oldukça fazla enerji gerektirmektedir. Mura ve ark. (1987), 1 mol pOrin halkası için
5
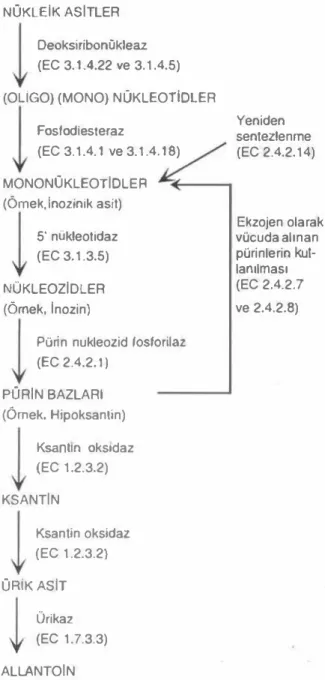
mol ATP'nin kul lanıldığını ileri sürmektedirler. Bütün pürinlerln ye niden sentezi, hipoksantinin nüklectidi olan Inazlnlk asit yoluyla meydana gelir. Bu metabolik yolun ilk adımı, glutaminin B- fosforibosilamin oluşturmak için losforibosil-1-plrofosfat (PRPP) ile reaksiyona girmesidir. Nükleik asiUer ve onların purin bazlan için enzimatik yıkıllmlar ve metabolik yollar Şekil 1 'de gösterilmektedir (Greife,1984).KAYA
Şekil 1. Nukleik asit (pGrin) senlezi, kaıabolizması ve ek· zojen olarak vOcuda alınan pürinlerin kullanılmasının bi·
yokimyasal yolları. NÜKLEIK ASITLER
i
Oeoksiribonükleaz
�
(EC 3.1.4.22 ve 3.1.4.5) (OLIGO) (MONO) NÜKlEOTIOLERi
Foslodieslerazı
(EC 3.1.4.1 ve 3.1.4.18)./
MONONÜKLEOT[OlEAK
(Ömek,lnozinik asil)i
5' nükleolidazt
(EC 3.1.3.5) NÜKlEOZ[OlER (Örnek, Inozin)i
PGrin nukleozidfosforilaz
�
(EC 2.4.2. i) PÜRIN BAZLARI ---' (Örnek, Hipoksanıin)i
Ksanlın oksıdazt
(EC 1.2.3.2) KSANTINi
Ksanıin oksidazt
(EC 1.2.3.2) ÜRiKASITi
""kaz-t
(EC 1.7.3.3) ALl.ANTOIN Yeniden senıezlenme (EC 2.4.2.14) Ekzojen olarak vücuda alınan pürinlerin kul· lanılması (EC 2.4.2.7 ve 2.4.2.8)Ekzojen olarak vücuda alınan punnlenn kul· lanımı, PRPP'nin punn bazlarıyla direkt reaksiyona girerek hipoksanlin fosforılNsil ıransferaz (EC 2.4.2.8) ve adenin fosforibasil transferaz (EC 2.4.2.7) kata1izörlü{ıünde guadinilik, inozinik, ksan· tilik ve adenilik asitlerin oluşumunu içerir (Murray 1971). O'Melto (1982) hipoksantin oluşumuna yön veren ksanlin oksidazın faaliyetini inhibe eden al· lopürinolün pürin salvage enzimlerini slimure etti{ıi, feed-back etkisini desteklediğini göstermiştir.
Idrarda Bulunan Pürin Oerlvatlarının Mikrobiyel Protein Senteziyle Olan Ilişkisi
Mikrobiyel proteinin ruminanta olan kaıkısı sa dece tüketilen yemin miktarı ile tespit edilir, fakat ru menin dolması ile ilişkili olan yemin fiziksel yapısı (hacim ve büyüklük) da bunu etkilemektedir. Alınan yemin miktarının azlığına bağlı olarak az miktarda digestanın geçişiyle sonuçlanan beslenme du rumlarında (Ozeilikle ruminal yıkımlanması fazla olan yemler verildi{ıinde) kaba yem ilavesiyle alınan mik tarın artırılması mikrobiyel protein akışını artırır_ (Chen ve ark. 1992a) Auminantlarda protein kul· lanımının elkilillğinin, rumen mikroorganizmalarının katabalik ve sentetik aktivitelerine ba{ılı oldu{ıu bi linmektedir. Etkilili{ıin fazla olması Için rasyon pro teini ve di{ıer nitrojen bileşiklerinin yıkılım Orün lerinin, rumende mikrobiyel proteine dOnüşümlennin hızlı olması gereklidir.
Pürin derivatları (PO) ço{ıunlukla rumende sen lezlenen mikrobiyel yapılardan kôken alırlar ve yem tüketim miktarını belirleyici bir niteliğe sahiptirler. Ayrıca
PO
atılım düzeyi, ruminanılarda Ozellikle pro tein ve ene�i metabolizmasına ba{ılı olarak değişen rumendekl mikrobiyel sentezlenmenin oranını da belirleyebilir (Mayes ve ark. 1995).Koyun ve sı{ıırlarda Ince bağırsaklara gelen nükleik asitlerin nel sindirilebilirliği sırasıyla yaklaşık ANA için % 80-90 ve DNA ıçın % 7S-8S'dir (McAI [an, 1982). Emilen pünnler, pürin kalabolizmasının ürünleri olan ve
PO
olarak tanımlanan hıpoksanıın, ksantin, ürik asit ve allantolne yıkımlanırlar. Yıkıltm ürünlerinin temelini aUantoin oluşıurur. PO ince ba ğırsaklardan emilen ve doku nükleik asıtlerinın dö nOşumünden açığa çıkan purlnler olmak üzere iki farklı kaynaktan kOken alırlar. Ruminantlarda ru· menden bağırsaklara geçen mikrobiyel yapılar ince ba{ıırsaklardan emilen pürin nükleotidlerinin temel kayna{ııdırlar (MeAlian ve Smith, 1973). PO idrarla atılırlar ve idrarla atıhm mikrobiyel kökenli pürin du zeylerlyle ilişkili oldu{ıundan ba{ıırsaklardan emilen mikrobiyel·protein düzeylerininde göstergesidir. id rardaki PO'nl Olçerek mikrobiyel protein katkısının belirlenmesi temeline dayanan teknik, hayvanların kanülünü gerektiren halihazırda kullanılan yön· temlere alternatif olarak kullanılan basit ve geçerlibir tekniktir (Chen ve ark. 1992a). Rumenden gelen nükleik asillerle, idrarla atılan pürin miktarı ara
sındaki ilişkiyi tanımlayabilmek için, aşa{ııdaki du rumlar göz Onüne alınmalıdır.
OeQişik yemlerne durumlarında rumen nükleik asitlerinin sindirim ve emiliminin devamlı etkilili{ji,
Endojen pOrinler üzerine gıda pürinlerinin ön leyici etkisinin eksikliği,
Endojen pOrin sentezi ve depo kaynaklarından kullanma yolları üzerine alınan ene�i seviyesinin et kisinin eksikll{ıi.
Idrana atılan diğer pürin rnetabolitlerine göre altantoinin oranının devamlılı{ıı (Giesecke ve arlı:, 1984).
Rasyondaki nükleik asitler rumende fazlasıyla yıkımlanırlar ve PD atılımına çok az katkıda bu lunurlar (McAllan.1980).ldrardaki PO'larının bazılan direkt olarak hayvanlann dokularından orijin alırlar ve ıoplam aııııma bu endojen katkı mutlaka gOz önüne alınmalıdır. Daha önceki bildirimlerde, bes lemede yapılan değişikliklerin, örn�in protein (Fu jihara ve ark.1987) yada enerjı Ilavesi gibi (Gi esecke ve arlı:, 1984), endojen PO'nın atılımı üzerine etkilerinin az oldu{ıu vurgulandı. Bununla birlikte de{ıişik lizyolojik durumlar1a ilişkili olarak en dojen kayıpların varyasyonları, duadonal ilave ve PO'nın idrarla atılımı arasındaki ilişkiyi tanımlamak amacıyla bu varyasyonların nasıl dikkate alınacağı hakkındaki bilgiler yetersizdir.
Endojen ve Ekzojen Pürinlerin Metabolizması Idrarla atılan endojen PO'I doku nükleik asit lennin dönüşümünden orijin alırlar. NOkleik asitlerin yıkilımından açığa çıkan pOrin nOkleozidleri ve ser besI bazların ço{ıu nOkleolidlerin yeniden sen tezlenmesi için kullanılırlar. Nükleik a·sil yı kımlanmasının ara ürünlerinin daha etkili kullanılabildi{ıl nOkleozid ve serbest bazlan içeren bir substrat siklusu vardır (Metzler. 1977). Ksantin oksidaz bu siklusta hipoksantini, ksantine ve daha sonra ürikaz varlı{ıında allantoine aksicte editebilen ürik aside dönüştürür. KsanUn. hipoksantin·guanln 'osloribasi! transferaz tarafından ksantin mo
nofosfat oluşturmak için yeniden kullanılsa da, ürLk
asit ve allantoin tekrar kullanıla;.maz (Gots, 1971).
Enzimin ksantine karşı aflinitesinin az olması ne deniyle bu reaksiyon yavaş olur (Hitchings,1978).
Bu nedenle ksantin oksidaı muhtemelen idrarda PO'larının kaybını tanımlayan anahtar enzimdir. Plazmanın aktivitesi basitçe di{ıer dokuların ak· tivitelerinl yansıımasına rağmen, sığır, koyun, domuz ve Insan arasında ekzoJen olarak vücuda ahnan pOrinlerin kullanımındaki larklılıkların ço{ıu, te orik olarak kandaki ksantin oksidaza atfeditebilir. Sığır, koyun, domuz arasındaki endojen PD'! atı lımının tür farklılıkları atılan derivatların yapılarıyls da tanımlanabilir. Ksant!n ve hipoksantin Iraksiyonları koyun ve domuzun idrarındaki toplam eks kresyonun kayda değer bır miktarını oluştururlar.
Hayvanların plazma ksantin oksidaz ak
tivitelerindeki farklılıklar nedeniyle, bunlar sığır id rarında pek kayda d�er görülmez. Sığır ve
do-muzların idranndaki allantoinin oranı
koyununkinden daha fazladır. Bu üç tür hayvanın kanında ürikaz olmadı{jı veya sadece iz miktarlarda bulunduğu için ürLk asidin allanloine dönüşümü dl{ıer dokularda meydana gelir. Bu nedenle doku Orikaıı aktivitesinin sı{ıır ve domuzlarda ko yunlardan daha fazla olduğu Maya çıkmaktadır (Chen ve arlı:. 1 990a).
Ruminantların normal fizyolojik koşullarda en dojen PO'nm ölçümü, rumen mikroorganizmalarının katkılarının ortadan kaldırılmasındaki teknik güç lükler nedeniyle engellenmiştir. Endojen atılımı ölç mek için, hayvanlar uzun sOre aç tutulmalarına ra{ı men, hayvanların metabolik aktiviteleri ya da doku nOkleik asitlerinin yıkımlanma oranları ve buna bağlı olarak pürin son Orunlerinin atılımının beslemedeki sınırlamayla değişebilece{ji ihtimali vardır. Bu prob lem inıagastrik Infüzyon tekni{ıinln kullanımıyla çö zülebilir (Orskov ve ark. , 979). Bu teknikte hay vanlar, uçucu ya{ı asillerinin rumene ve kazenln de abomazuma Inmze editmesiyle beslenirler. Ru mendeki mIkrobiyel fermentasyon elimine edilir ve hayvanların normal besleme şar1ları bu yöntemle muhafaza edilebilir.
Belli bır sınıra kadar rasyana yapılan ilaveler endojen PO ekskresyonunu etkileyebilir (Giesecke ve arlı:. 1984, Lindberg ve ark 1989. Chen ve arlı:. 1992b). Endojen pOrin derivatan olan allantoın, Ori kasit ve ksantin+hipoksanlinin idrarla atılımını ölç mek amacıyla, koyunlar ve yetişkin sığırlar Int ragastrik inmzyonla, buzağılar süt ikame yemleri ve proteinsiz rasyonlarla beslendiklerinde, toplam
KAYA
PO'larının sı!:j:ırda (514 (SE 20.6) mmollkg WO·151 gün), koyun (168 (SE
5.0)
mmoV kg WO·15/g0n) ve domuzla (166 (SE 2.6) mmoV kg WO·15/gün) kar şılaştırıldığı zaman Onemli miktarda fazla olduğu görülmüştCir. Sığır plazması ksanlin oksidaz (EC 1.2.3.2) aktivitesine sahip olmasına rağmen, koyun ve domuz plazmasının bu aktiviteye sahıp olmadığı tespit edilmiş, Orikazın ise slQır ve domuz plaz masında (EC 1.7.3.3) bulunmadı!:j:ı, koyun plaz masında iz miktarlarda olduğu bildirilmiştir. Bu nedenle endojen PO atılımındaki tOr farklılıklarının, dokulardaki ve özellikle kandaki ksanlln oksidaz aktivitesinin farklı profillerine ba!:j:lı olarak, yüksek ksanlin oksidaz aktivitesinin pı1rinlerln yeniden kul lanılamayan bileşiklere yıkımlanmasını artırmasıyla meydana geldiQi ifade edilmektedir (Chen ve ark.199Oc). Rasyon proteini nOkleik asil lçermediği zaman. hipaksanıin, ksanlin ve Orik asitin al lanloıne dönuşOmOnden sonra endojen allanloinin atılımı sığırlarda ve kuzularda sırasıyla 72 ve 26 mg/kgWO·15/g0n olarak hesaplanmıştır (Fujihara ve ark. 1987).Dişi domuzlarda gebeliğin endojen PO ekskresyonunu elkilemediQl, bu nedenle idrardaki PD'nın atılımından pı1rinlerin duedonuma ge çişleriniı değerlendirmek amacıyla daha önceden gebe olmayan hayvanlar için geliştirilen (BalceJls ve ark. 1991) denklemlerin gebe hayvanlarla uzun sureli kuJlanımlarda geçerli olmadlQı be lirtilmektedir (Martin Orue ve ark.199S).
BalceJls ve ark.(1993), n[!roJen ilavesiyle top lam PO'nın idrardaki atılımında artış oldu{ıunu ve bu cevabın atılan allantoinin artmasına bağlı olarak (26.9-66.4 mglkgWO·15/g0n) meydana geldiğini öne sürdüler. Ksantln, hipoksantin ve urik asit atı lım oranlarının sırasıyla ortalama 1.8 (SE 0.17), 5.4 (SE 0.21) ve 7.2 (SE 0.36) mg/kgWO·75/gün olduğu ve fazlaca değişiklik göstermediği bildirildi. Mi nimum amonyak konsantrasyonunun (yaklaşık
50
mgIL) muhafaza edilmesinin, kuru madde alımı ve fermentasyon oranındaki anlamlı azalmalardan sa kınmak için gerekli oldu!)u, bununla birlikte ru mendeki NHa konsantrasyonunun belli bir oranın Ozerine çıkmasının bir sonucu plarak idrarta atılan allantoinin ve duedonumdaki mikrobiyel N akımının artabildiği ılade edilmektedir.Diğer sınırlamalar olmadı!)ında, mikrobiyel enerjetik etkililik olarak tanımlanabilen fermente edilmiş enerjı ilavesi ve rumenden gelen mikrobiyel ürOn arasındaki itişkinin, toplam sentezlenmeye karşı mikrobiyel OrOn akışının oranı ve mik roorganizma populasyonunun enerji harcamalarının karşılanması ile etkilendi§i bildirilmektedir. (Oew hurst ve Webster, 1992). Oewhurst ve ark. (1987), enerji ilavesine ilişkili olarak, idrarla aUlan allanloin miktarının rumenden gelen solidlerin fraksiyonel akış oranındaki (Ks) değişikliklerle etkilendiğini or taya koymuşlardır.
Koyunlar için sabit endojen kalkı olduğu dü şunulerek yapılan çalışmalarda ise, tespit edilen geri elde etme de{ıerleri değişkenlik
(0.15-1.00)
arzetmiştir (Tabıoı). Yapılan araştırmalar emilen pOrinlerin koyunlarda doku nOkleik asitlerine dö nOşebildiğinl göstermektedir (Condon ve ark. 1970, Smith ve ark. 1974, Razzaque ve ark. 1981). Bu nedenle emilen pürin miktarına göre PO'nın idrarla atılım oranı linear deQlldir. Ancak sı{ıırlarda sınırlı da olsa emııen pOrlnlarin dokularda kullanımı söz konusu olduğundan dolayı bu oran lineardır (Verbic ve arı<. 1990 Tablo 1).Ekzojen pOnnlerin Idrardaki PO'na katkısı, purin nukleotidlerinin, ANA ya da izole bakterilerin post ruminal InfOzyonuyla tespit edildi (Mayer ve ark. 1995). Ekzolen pı1rinler, pürin nı1kleozidleri ve serbest bazlan olarak bağırsaklardan emilirler (Wil son ve Wilson 1962). E!)er bağırsak duvarını ge çişlerinde ksanlin, Orikasi! ve allatoine dö nOşmezlerse, ekzojen olarak vucuda alınan pOrinlerin doku nOklelk asitlerinin oluşumunda kul lanılmaları söz konusu olacakıır (ksanlfn çok düşOk bir oranda yeniden kullanılabilir) (Hitchings, 1978). Sığırdakinin aksine, koyun intesıinal mukozası çok az da olsa ksantin oksidaz aktivitesine sahiptir (Ak halidi ve Chaglassian, 1965) ve bu nedenle ekzojen pürinlerin yeniden kullanılamayan derivatlara dö nüşümO anlamlı bir miktar teşkil etmez (Chen ve ark. 1990b).
Şayet bağırsak dokusundan emllen pürinlerin yıkımlanması anlamlıysa, portal dolaşımdaki urik asit ve aUantoinin konsantrasyonları fazla miktarda olmahdır. Bu nellenle, koyunlarda ba!)ırsak ve ka raciğer dokularına giren emilmiş pürinlerin çoğunun
Tablo 1. Koyun ve sığırda infUze edilen pOrinler ya da nOkleik asitlerin geri ekie edilmesi.
Infüzyon B61gesl Substrat Geri Etde Edilme Kaynaldar
Koyun
Abomasum (14C) Adenin 32 Condon ve ark 1970
Abomasum ANA 75-90 Condon ve Hatfield 1970
Rumen Işaretli bakteri 15 Smith ve at1(. 1974
Duedonum ANA 29 Antoniewicz ve ark. 1980
Rumen Işaretli bakteri 20·27 Razzaque ve at1(.1981
Duedonum ANA 52 Giesecke ve ark. 1984
Abomasum Pruten' 82·100 Fujihara ve ark.I987
Abomasum Pruten n Chen ve ark.l99Ob
Duedonum Digesta+RNA 80 Balcalls ve ark. 1991
IntravenOz Aııantoin 72 Chen ve at1(.1991
Sı?)ır
Ouedonum Adenin 65 MeAlian, 1980
Abomasum Pruten 77 Verbic ve ark. 1990
Pruten: Proleln konsantrasyonu olarak kullanılan metanal substralından elde edilen bir bakteri kültürOdür. kullanılabilir larmlarda oldu�u belirtilmektedir.
(Chen ve ark. 1990b).
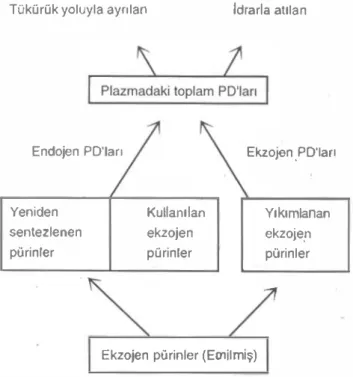
PO vücutta farl<lt enzim profillerine sahip farl<lı bölgelerde üretilir1er. Koyunda rumen bakterileri nOkleik aistlerinin kullanımı ölçüldü�ünde, en az %5'inin karaciOer, dalak ve böbrekte bUlunduOu, di�er %20'sininde kaslarda görüldüOü ifade edil miştir (Smith ve ark. 1974). Endojen PO'ları tüm do kularda üretilir. Koyunlarda tüm ekstrahepatik hüc· reler çok az sevide ksantin oksldaz aktivitesi içerirler(AI·Khalidi-Chaglassian,I965). Bu enzimin etkisiyle ksantin, hipoksanıin ve ürik asit olarak kana geçen ve orada ıaşınan endojen PO'larının oranları fazla olmaktadır (Chen ve arı<. 1990<:). Bu· nunla birlikte ekzojen pOrinlerin yıkılımından kökan alan PO, ksantin oksidaz ve ürikazın yüksek ak tivitelerine sahip olan karaciOerde meydana getirilir ve PO'nın ço{ıu allanloin formundadır. Bu nedenle, pıazmadaki ve dolayısıyla idrardaki PO'nın son kompozisyonu direkt olarak endojen ve ekıojen kaynaklardan gelen katkıya göre deOişmektedir. Koyundaki ekzojen pürinlerin emilimi ve PO'lerinin idrar1a atılımı arasındaki ilişki Şekil 2'de gös terilmektedir.
Emllen mIkrobiyel pürinlerin-vücuda katkısı az az oldu�u zaman, PO atılımının emilen mikrobiyel
purin miktannı ölçmede kullanılabilmesI için, idrar1a atılan PO'nın toplamına etkı eden endojen katkıyı belirlemek amacıyla bir düzelimeye ihtiyaç olacaktır. Bu dOzeitme idrardaki toplam atılım 0.6 mmoV kgWO·75/gün'den daha az olduOu zaman uy gulanmaktadır (Chen ve arı<. 1990b).
Y=Ü.84X+ (1.50BWO·75 e -O.25x )
Y= Endojen ve ekzojen orijinli toplam PO'larının idrardaki atılım miktarı (mmoVgün), X", Emilen mik robiyel pünnlerin miktarı, 1I'JO.75 = Metabolik vücut a�lr1r�1. Bu model, parantez içensinde sunulan şek· liyle, ekzojen pürinlerin azalmasıyla hayvanlar la· rafından kullanım için hazır olan endojen pünnlenn katkısını doOru olarak belir1emede kullanılır. Ek Iemdeki Y'den X'in hesaplanmasının Newton'un ite ralien işleminin ortalamasıyla yapıldığı ifade edil· mektedir(Chen ve ark.1992a).
Idrardaki toplam PO atılımı yaklaşık 0.6 mmoV kgWO·75/gün'den daha fazlaysa endojen kalkının çok az olaca�ı ve sıfır olarak alınabileceği bil dirilmektedir. Genellikle , enerji ihliyaçlarının yak laşık 0.8 katı ve Ozerinde normal olarak beslenen koyunlara bu uygulandlOında, emilen pünnlerin mik larının aşağıdaki� formüle göre hesaplanması ge rektiOl ifade edilmektedir.
KAYA
X=d 10.84
x= Emilen pürinlerin mikıarı, d= Toplam PO'nın idrardaki aıılımı (Chen ve ark. 1990b).
Pürinlerin giriş-çıkış ilişkisine bakılarak, kana giren PO'nın koyunlarda (n=6) %84'ü, sı�ırlarda (n=2) %85'inin devamlı olarak idrarla alıldı�ı gös leriidi. Allanloin ve ürik asitin %10 kadarının lü kürük yoluyla geri döndükleri ve orada lamamen yı kımlandıklan bitdirildi (Chen ve ark. 199Oc). Glomeruler filırasyon oranı de�işmedi�i için, al lanloinin plazma konsanırasyonu direkt olarak en dojen allanloinlerin girişiyle ölçülebilmektedir. Gre ger ve ark. (1976), ratlarda nefron boyunca allanıoinin geri emilimi ya da sekresyonunun 01-madı�ını ileri sürdüler. Glomeruler fillraldat) al lanıoinin nel geri emiliminin koyunda meydana gel di{JI, fakat kapasitesi sınırlı oldu�undan endojen allanloin üretimine eşil (- 2.9 mmol/gün) tubuler ka pasiteyle doygunlaşıırıldığı ifade edilmektedir. Bu nedenle kana herhangi miktar allantoin ilavesinin glomeruluslar içine filtre edildikten sonra idrarda atılacağı bildirilmektedir (Chen ve ark.1991). Şekil 2. Koyundaki ekzojen pürinlerin emilimi ve
PO'lerinin idrarla atılımı arasındaki ilişki (Chen ve ark. 1990b).
TOkOrOk yoluyla ayrılan Idrarla atılan
Endojen PO'ları Ekzojen.PO'ları
Yeniden Kullanılan Yıkımlanan sentezlanan ekzojen ekzojerı
pOrinlar pOrinler pürinler
Ekzojan pOrinler (Emilmiş)
Koyunlar, allantoinin fizyolojik miktarlarını ölç mek için uçucu ya{J asitleri ve kazeinin intragastrik infüzyonuyla beslendiklerinde, geri alınımın % 72 oldu�u ve geri kalan kısmın bağırsak içine diffüze olduğu ve bağırsak mikroflorası tarafından yı kımlanması ile kaybolduğu bildirilmektedir. Kre atinine dayalı hesaplamada, allanloinin glomeruler filtrasyon oranının ve tubuler geri emiliminin inl ravenöz infüzyonla değişmeyeceği gösterildi. Gün lük maksimum 1.28 mmol olan tubuler geri emilim, tek başına endojen al1antoinin yüklenmesiyle doy gunluğa ulaştırıldl. Yapılan araştırmada, koyunlar normal olarak beslendiğinde allantoinin atılımında iki katı farklılık olduğu, sindirilen her ünite yem için hayvana olan mikrobiyel katkının yemlerne se viyesiyle etkilenebildiği ifade edilmektedir (Chen ve arı<. 1991)
Idrarla atılımın yanı sıra, PO sütle atılabilir veya lükürük yoluyla kaybolabilir. Kirchgessner ve Kra uzel (1985) toplam pürin derivallarının yaklaşık 0.06-0.07'sinin süt yoluyla atıldığını göstermişlerdir.
Idrardaki ve sütteki allantoinin endojen pürinler kadar rumenden de kaynaklandığını düşünmek ge rekir. Pürin metabolitleri olan allanloin ve ürik asit sütçü sl{Jırlarda sadece idrarda değil kayda deOer miktarlarda sütte de tespit edilmektedir. Sül üre timine bağlı olarak değişen sülteki allantainin mik tarıı toplam allanloin atılımının % 1-4 arasında ol masına rağmen; bu alııımın, rumendeki mikrobiyel gelişimin temel gösıergesi olan laktasyandaki nel enerji alımıyla (NEOolan ilişksii idrardan ziyade süt için daha yüksektir. Holstein sı�ırlarda idrarda ve sülteki allantainin atılım miktarı sırasıyla yaklaşık 294 ve 4.1 mmoVgün iken ürik asil için bu de�erler 35 ile 1.1 mmollgün olarak de{ıişliği bildirilmektedir (Giesecke ve ark.1994).
idrardakl Pürin Derivatlannın Yem Tüketiminin Tespitinde Kullanımı
Rumendeki mikrobiyel sentez ene�i ba{Jımlı ve fermenle edilebilir substratların elde edi lebilmesiyle yakından ilişkili olduğu için, PD'nın atı· Iımı rumende fermente edilebilir organik maddelerin alınımını yansıtmaktadır. EOer sindirimin ruminal ve poslruminal kısımları bilinseydi, PD'larının atılımı sin dirilebilir organik madde (OOM) alınımının bır gös tergesi olarak kullanılabilirdi. Değişik ruminanılarda yapılan çalışmalarda, allantoinin idrar1a atılımının
DGM alımıyla pozitif bir ilişkiye sahip oldu!:i'u gö.
teriidi (Vercoe, 1976, Antoniewlcz ve ark, 1981,
Lindberg, 1985, Balcells ve ark. 1993, Liang ve
ark. 1994). PD'nın atılımıyla DOM alımı arasındaki
ilişki, fermenle edilen her ünite maddeye göre du
edonuma giren mikrobiyel protein miktarının de
ğişmesi gibi mikrobiyel proıeln katkısını etkileyen
faktörlerle de!:i'işebilmektedir. Yemlerne seviyesinin
rumendeki mikrobiyel sentez üzerine etkisini araş·
tıran Chen ve ark.(1992a), PO atılımını mikrobiyel
ürünün bir göstergesi olarak kullandıklarında,
DGM'nin her ünitesi için oluşan mikrobiyel azotun,
her bir yaşam ağırlığı için tüketilen yem miktarıyla
ilişkili olduğunu gözlediler. Bu aynı rasyon için bile
PD:OOM orantısında iki katına ulaşabilecek fark
lılıkların beklenebildiğini ifade etmektedir. Buradan
daha da ileri gidilerek mikrobiyel ürünün ve buna
bağlı olarak PO atılımının yem tüketiminin linear bir
fonksiyonu olmadığı ortaya çıkmaktadır.
Tüm bu gözlemler, PO atılımının şayet belir1i
rasyonlar1a beslenen belirli ruminantlar Için özel bir
ilişki tespit edilmişse,o zaman yem tüketiminin bir
göstergesU olabileceğini öne sürmektedir. Böylesi
bir uygulamanın meradaki hayvanlar için dü
şünülebilecegi ifade edilmektedir (Mayes ve ark.
ı
995).
Sığır ve koyunlarda, mikrobiyel protein akım
miktarını belirlemek için yapılan araştırmalarda
(Chen ve ark. 1990b, Verbic ve ark.1990 ve Bal
cells ve ark. 1991) değişik modeller geliştiriidi.
Ancak
bu
çalışmalarda,
rumen
mik
roorganizmalarındaki pürin
N :Toplam
Noranı ve
emilen pürinlerin geri elde edilmesi, tanımlanmaları
gerekli olan önemli iki parametreyi oluşturmaktadır.
Mikrobiyel proteinin bağırsaktaki akışını tespit
etmek için uygulanan PO atılımı ve diğer metotlar
(35S ya da 15
Nye dayalı) arasında bir kar
şılaştırma yapılmış ve her bir çalışma içerisinde
(Lindberg ve ark. 1989, Puchala ve Kulaseck,
1992, Perez ve ark. 1994) PO atılımı ve mikrobiyel
Nakışı arasında karşılıklı bir ilişki olduğu fakat de
neyler arasında önemli farklıtıklar bulundu�u so
nucuna varılmıştırr. Bunların duedonuma giren mik
robiyel materyaldeki pürin
N;toplam
Noranındaki
ya da emilen pürinlerin idrarda elde edilmesindeki
farklılıklara bağlı olarak meydana gelebildiği ifade
edilmektedir (Mayes ve al1<.1995). Bu be
lirsizliklerin nedeninin bilinmesi gerekli olmakla bir
likte, ruminantlarda mikrobiyel protein üretimi ve
onu etkileyen faktMer üzerinde çalışmak için idrar1a
atılan PO'nın Olçümü metodu, çok elverişli ve basit
bir metot olarak sunulmaktadır.
Kaynaklar
A1-Khalidi, U.A.S. and Chaglasslan, T.H. (1965). The
SP'!
des Dısıribuliorı ol Xanıhine Oxidase. Biochem.J., 97, 318-320.Antooiewicz,A.M., Heineman, W.W. and Hanks, E.M. (1960). The Elleel ol Changes in the Intestınal Ftow ol Nuc· reic Aclds on AltanIcin Excrelion in the Urine ol Sheep. J.Agrk:. ScI.,Camb.,95, 395-400.
Anıoniewicz,A.M., Heineman, W.W. and Hanks, E.M. (1981). Ellecı ollevel ol Feed Inlake and Body Masson on Allantcin Excreıion and lhe Allanıcin ıo Creaıinine Raıio in the Urine ol Sheep. Rocznikl Naukowe Zootechnikl H.,8z, 1, 49-65. Salcells, J., Guada. J.A., Gasırlllo, C. and Gasa. J. (1991).Urirıary Excretion ol AIlanlOln and AIlanıcin Pre
CUI"SOl'S aher Dıfferent Raıes of Punne Inlusion inlo lhe DlI edonum. J., Agric. Sci .. C8mb .. 116, 309-317.
Balcells, J.. Fondevila, M., Guada, J.A., Castrillo, C., and Surra, J.C.E.(1993). Urinary Excrelions ol Punne Derivalıves and Nıırogen in Sheep Givens Slraw Supplemenled wilh Dif ferenl Sources of Carbohydraıes. Anlm. Prod., 57, 287-292. Chen, X.B., Chen, YK, Franklin, M.F., Orskov, E.R. and Shand, W.J. (1992a). The Eflecı of Feed Inlake and Body Weighl on Purine Darivaıive Exceılon and Microbial Prolein Supply in Sheep. J.Anim.Sci.,70, 1534-1542.
Ch&n, X.B., Chowdry, S.A., Hovell, F.D. DeB., Ors/(;ov, E.R. and Kyle, o.J. (1992b).Endogenous Altanıein Excretioo in Response lO Changes in Proıein Supply in Sheep.J. Nulr.,I22,2226·2232.
Chen, X.B., Havell, F.o. oeB. and Orskoy, E.R. (199Oa).Exctetiorı ol Purine Derivalıves by Ruminanıs: See relion ol AltanIcin imo the Rumen via Saliva and Its Faıe in the Gut. Br. J. Nutt.,63,197-205.
Chen, X.B., Havelt, F.o. DeB., Orskav, E.R.and Brown, D.S. (199Ob). Excrelion of Purine Derivalıves by Ruminanıs: EI lects ol Exogenous Nucleic Acid Supply on Purine Oerivative Excretlon by Sheep. Br. J. NUlr., 63,131-142.
Chen, X.B., Kyte, O.J., Orskov, E.R. and Hovetl, F.D. OeD. (1991).Renal Clearenee ol Plasma Allanıcin in Sheep. Exp. Physioi., 76, 59-65.
Chen, x.a., Orskoy, E.A. and Hovell, F.o. DeB. (199Oc).Excretlon of Purine Derivatives by Ruminants: En
dogenous Excretion, Dillerences betweenCattle and Sheep. Br. J. Nuır., 63,121-129.
Condon. R.J., Hall, G., and Hatlield, E.E. (1970). Metabolism of AbomasaJly Infused 14C-labeJled RNA Adenine, Uracil and Glycine.J. Anim Sci.,31,1037-' 038(Absır.)
Coodon, R.J. and Hatf18ld, E.E. (1970).Meıabolism ol Abc> masally Infusad Riborıucleic Acid by Sheep. J. Anim. Sci.,3ı, 1037A.
KAYA
Use ol Urlnary Allanıoin Excreıion ıo Assess the Energeıic Ellideney of Mıcrobial Protein Yield in Sheep. Anim. Prod., 44. 475A.
Dewhursı. R.J. and Websıer. A.J.F.(1992). EHeet 01 Diet, Lavet ol Intake, Sodıum Bicarbonale and Monensin on Uri· nary Allanloin Excfelion in Sh&ep. Br. J. Nutr., 67, 345·353. D'Mello. J.p. (1982). Ulilization of Dietary Purines and Pyri mldines by Non-Ruminanl Animals. Proc. Nutr. Soc.,41
(3).301-306.
Fuııhara, T" Orskov. E.r., Reeds, P:J. And Kyfe, D.J. (1987). The ellect of Prolell'l Inluslon on Unnary ExCfetioo of Purina DerivatlV9s in Auminanıs Nourished by tnlragasıric Nuı nllOl1. J.Agnc. Sa., Gamb., 109. 7-12.
Glesed<e , D., Ehrentreich, L. and Sıangassinger, M. (1994). Mammary and Renal EXCfeliorı of Purine Me ıabolites In Aetalion ıo Energy lntake and Milk Yield in Dairy Cows. J. Dairy. Sel., 77(8)2376-2381.
Giesecke, D., Sıangassinger, M .• r18meyer, W. (1984). Nuc
leic Acid Digestion and Ulinary Purine Metabolites. In (Sh&ep Nourished by Intragasıric Nutritıon, Gan. J. Anim. Sci., 64,suppt,144-145.
Gots, J.S.(1971). Reglllation of Punne and Pyrimidine Me tabolısm. In (Metaboli<: Aegutation, voI.5, pp.225-255.(H.J. Vogeı, ediıor). New York and London:Acad8mlc Press. Greger, R., Lang, F.and Deetjen, P.(1976). Renat excreıion ol Purina Metabolites,Uraıa and Ananlo/n. by the Mam· malian Kidney. In Intemational Review ol Physlology: KId ney and Urinary Traet Physiology tl, vo1.11, ad. K. Thurau, pp.257-281. University Park Press, Banimore, London, Tokyo.
Greile, HA (1984). Nutritive value of Dieıary Ribonucleic Acid and Jts Nucleosides lor Growing Rats with a Dieı DeL· licienı in Nonessenlial Amlno Acid Staıus ol Rats. Zentrabl Veterınarmed. A .. 31(4), 269-279
Hitchings, G.H.(1978). Uric Acid: Chemlsıry and Synlhesls. In Uric Acid. pp.I·lO (editors, W.N. Kelly and J.M. Weiner), Benin. Heidelberg and New YOrk:Springer-Venag.
Kirchgessner, M. And Kreuzer, M. (1985). Hamstolf und Ai· lanloln in der Miich vorı KuhenWahrend und nach Ver ıünen.ıng zu Hoher und zu Nıedrlger Protein·mengen. 5. Zum EinlluB von Proteinlehlfımahn.ıng bel Laktierenden Kühen und Daraus Enlstehendan Nebenwirl<ungen. J. Anim. Physiol.Anım. Nutr.,54, 141.
Uang, J B., Matsumo, M. And Young. B.A.(I994). Purine Derivabve Excretion and Ruminal Mlcroblal Yield in Ma laysian Canla and Swamp Buffalo. Anim. Feed. Sei. Tech.,47.189-199.
Undberg, J.E. (1985). Urinary AJianlain Excretion and Di geshble Organlc Matter Intake In Dalry Goats. Sweel. J. Agric. Res .. 15,31·37.
Undberg, J.E., Bristav, H. And Manyenga, A.R (1989). Exc retion of Purines in the Urine of Sheep in Relation ıo Du ecIooal F10w of Mlcroblal Proıeın. Swed. J. Agric.Res., 19, 45-52.
Martin On.ıe, S.M., Balcells,J .• Guada, J.A.nad Gastriıfo, C.
(1995). Endogenous Purine and Pyrimldine Oerivative
Exe-retion in Pregnant Sows. Br. J. Nutr.,73, 375-385
Mayes, R.w., Dove, H., Chen, X.B. and Guada, JA (1995). Advences in the Use of Fecal and Urinary Markers lor Me asuring Diet Compositlon, Herbage Intake and Nuırient Uli IIzallon In Herblvores. In "Recenl Developments in Ihe Nut rition of Herblvores Proceedlngs ol the Ivlh Intemational Symposlum on the Nuırillon ol Herblvores", Ed. M.Joumet, E.Grenel, M.H.Farce, M.Therlez. C. Damarquilly. pp.381-406,INRA Editions,Paris.
McAllan, A.B. (1980).The Degradation ol NucleicAcids in lhe n.men and Lhe Removal ol Breakdown Products Irom the Sman Intestines ol Steers. Br. J. NUlr.,44,99-112.
McAIIan, A.B. (1982).The Fata ol Nuclalc Acids in Ru minants, Proc. NUlr.Soc.,41,309.
McAJlan. A.B. and Smith, R.H. (1973). Degradalion ol Nuc leic Acid Derivatives by Rumen Baeteria inVilro. Br. J.Nutr.,29,467·474.
Metzler, B.E.(1977). Blochemistry: The Chemical Reactlons
of UVIrıg Cells,p.881. London:Academlc Press.
Mura, U., Osman, A.M .• MohamecI, A.S., Di Menino, D., and lpata, P.L (1987). Purine Salvage as Metabolite and Energy Saving Mechanlsm In Camelus dromedarius: The Recovery of Guanlne.Comp. Biochem. Physiol. B.,87(1),157-16O. MUrT8y, A W. (1971). The BioIoglcal Signlficance of Purine Salvage. Ann. Rev. Biochem., 40, 811-826.
Orskov, E.R" Gn.ıbb. DA, Wenham, G. And CorrigaJl, W. (1979). The Fastenance ol Growing and Fanenıng Ru mlnants by Imragasıric Inlusion of VOIatile Fatty Acids aOO Protein. Br. J. Nutr., 41, 553-558.
Perez, J.F., Balcells, J., guada, J.A. and Castnla, C.(I994). Mlcroblat·N Synlhesis in the Rumen:Urinary Purine De rivatives vs. 15 N as an Esıimative Methods, Proc. Soc. Nutr.
Physiol., 3, 233.
Puchala, R. And Kulased<, G.W. (1992). Esıimation ol Mit· roblal Protein FIow lrom the Auman ol Sheep Usıng Mic roblal Nuclelc Acid and Urinaıy Excretion ol Purine De· rivatlves., Can. J. Anim. $ci., 72, 821·830.
Razzaque, MA, Topps, J.h .. Kay, RN.B. and Brockway. J.M. (1981). Meıabolism ol the Nuclelc Acids of Rumen Bac terla by Pre n.ıminant and Ruminam Lambs.Br. J. Nulr., 45, 517·522.
Smith, R.C., Moussa, N.M. and Hawkins, G.E. (1974). Uti· lizatk>n of the Nudeic Acids ol Escherichia CQ{i aOO Rumen Baeteria by Sheep. J. BloI. Chem.,32.529-537.
Verbic, J., Chen, X.B., MacLoad, NA and Orskov, E.R (1990). Excretion ol Purine Derivatives by Ruminants: EI· lects of Mlcroblal Nuclelc Add Inlusion on Purlne Derivative Excreıion in Steers. J. Agric. $ci.,Camb. 114. 243-248. Verece, J.E. (1976). Urinary Aliantoin Excreıion and Di· gestible Dry-Maner Intake in Canle and Bullalo. J.Agric. Sci .. Camb, 86, 613-615.
WilSon, D.W. and Wilson, H.C.(1962). Studies in Vitro of the Digestıon and Absorptiorı of Purine Ribonudeotides by In testine. J. BioI. Cham .• 237, 1643-1647.
Zöllner, N. (1982). Purirıe and Pyrimldine Metabolism. Proc. Nutr.soc .• 41,329-342.