Metabolite Profiling by Hyphenated Liquid Chromatographic Mass
Spectrometric Technique (HPLC-DAD-ESI-Q-TOF-MS/MS) and
Neurobiological Potential of Haplophyllum sahinii and H. vulcanicum
Extracts
Esengul Karahisar,
aOsman Tugay,
bIlkay Erdogan Orhan,*
cFatma Sezer Senol Deniz,
cSimon Vlad Luca,
d, eKrystyna Skalicka-Wozniak,*
dand Mustafa Sahin
faDepartment of Biology, Faculty of Science, Selçuk University, Konya 42250, Turkey
bDepartment of Pharmaceutical Botany, Faculty of Pharmacy, Selçuk University, Konya 42250, Turkey
c
Department of Pharmacognosy, Faculty of Pharmacy, Gazi University, 06330 Ankara, Turkey, e-mail: [email protected]
dDepartment of Pharmacognosy with Medicinal Plant Unit, Medical University of Lublin, 20-093 Lublin, Poland,
e-mail: [email protected]
eDepartment of Pharmacognosy, Faculty of Pharmacy, GrigoreT.PopaUniversity of Medicine and Pharmacy Iasi,
700115 Iasi, Romania
f
Department of General Surgery, Faculty of Medicine, Selçuk University, Konya 42250, Turkey
In the current study, the ethanol extracts of flower, stem, and root parts of two endemic Turkish species, e. g.,
Haplophyllum sahinii O. TUGAY & D. ULUKUŞ and H. vulcanicum BOISS. & HELDR., were screened against
acetylcholinesterase (AChE) and butyrylcholinesterase (BChE) associated with Alzheimer’s disease as well as tyrosinase (TYR) linked to Parkinson’s disease using ELISA microplate assay at 200 μg/mL. Among the extracts,
the highest inhibition was caused by the stem extract of H. sahinii against BChE (IC50=64.93 � 1.38 μg/mL).
Consistently, all of the extracts were found to exert a selective inhibition towards BChE to some extent. It was
only the root extract of H. vulcanicum that could inhibit AChE at low level (IC50=203.18 � 5.33 μg/mL). None of
the extracts displayed an inhibition over 50 % against TYR. Metabolite profiling of the extracts was achieved by a highly hyphenated liquid chromatographic mass spectrometric technique (HPLC-DAD-ESI-Q-TOF-MS/MS), which revealed the presence of furoquinoline (β-fagarine, γ-fagarine) and amide (tubasenicine, tubacetine) alkaloids; furano- (rutamarin), pyrano- (xanthyletine), and geranyloxy coumarins; phenylpropanoid (secoisolariciresinol), arylnaphthalene (mono-O-acetyldiphyllin apioside), and dibenzylbutyrolactone (kusunokinin, haplomyrfolin) lignans. Several important differences were observed between the extracts analyzed. β-Fagarine was the major alkaloid in H. vulcanicum, whereas γ-fagarine was present only in the roots of both Haplophyllum species; moreover, secoisolariciresinol and secoisolariciresinol dimethyl ether were the main lignans in the stems and flowers. This is the first study identifying ChE and TYR inhibitory effect and metabolic profiles of H. vulcanicum and H. sahinii.
Keywords: Haplophyllum sahinii, Haplophyllum vulcanicum, cholinesterase, tyrosinase, metabolite profiling, biological activity.
Introduction
The genus Haplophyllum A.JUSS(Rutaceae) comprises a
total of 19 taxa, of which 11 are endemic to Turkey.[1,2]
Haplophyllum species are amongst important
medici-nal plants and some of them, e. g., H. telephioides BOISS.,
were reported to be used in Anatolian folk medicine,[2]
while many species of this genus are also used in traditional medicine for the treatment of injuries, ulcers, diabetes, infertility, fever, menstrual pain, diarrhea, hypertension, and skin diseases in various
countries. Furthermore, numerous pharmacological activities such as antioxidant, antimicrobial, insectici-dal, antiprotozoal, mollusciciinsectici-dal, cytotoxic,
cardiovas-cular, anti-inflammatory, and acetylcholinesterase
(AChE) inhibitory effects, have been reported for the
genus.[5] Diversity in phytochemical content of
Hap-lophyllum species has been so far confirmed by the
presence of mainly alkaloids, lignans, coumarins, and
flavonoids.[6]
At the moment, cholinesterase inhibitors (ChEIs) are considered as the most effective drug class for the treatment of Alzheimer’s disease (AD), which is a degenerative and progressive neurological disorder characterized by memory and thinking impairment
and abnormal behaviors.[7,8]Although the
pathogene-sis of the disease has not been fully explained due to its multifactorial character, one of the most accepted theories is the ‘cholinergic hypothesis’, which is defined as a deficiency in the levels of the neuro-mediators acetylcholine (ACh) and butyrylcholine (BCh) observed in the brains of AD patients.
Con-sequently, the inhibition of acetylcholinesterase
(AChE) along with butyrylcholinesterase (BChE), the key enzymes which hydrolyze ACh and BCh, respec-tively, has become a major treatment option towards
AD.[9] Additionally, ChEIs have been also used in the
treatment of glaucoma and myasthenia gravis or as
insecticidal agents.[10 –12]On the other hand, tyrosinase
(TYR), an enzyme found in melanocytes, catalyzes the rate-limiting oxidation of tyrosine to melanin, which plays a critical role in the pigmentation of skin, hair, and eyes in animal, undesirable browning of fruits and vegetables, moulting process of insects, and dopamine toxicity in Parkinson’s disease (PD) as well as neuronal
death in PD and Huntington’s diseases.[13,14] Thus,
inhibition of TYR might be a new and quite important target in the treatment of the aforementioned dis-eases.
In the present work, we aimed to investigate inhibitory potential of the ethanol extracts prepared from the flowers, roots, and stems of H. sahinii and H.
vulcanicum, two endemic species to Turkey, against
AChE, BChE, and TYR enzymes. Besides, the extracts were subjected to phytochemical profiling by a highly hyphenated liquid chromatographic mass spectromet-ric technique (HPLC-DAD-ESI-Q-TOF-MS/MS).
Results and Discussion
HPLC-DAD-ESI-Q-TOF-MS/MS Metabolite Profiling
Alkaloids, lignans, coumarins, and flavonoids are considered among the most significant chemical markers of plants belonging to Haplophyllum genus with more than 75 different molecular entities
reported.[6] While H. sahinii was not previously
inves-tigated, H. vulcanicum has afforded up to date alkaloids (vulcanine, dicatamnine, skimmianine, robus-tine, haplopine, γ-fagarine), lignans (haplomyrfolin, kusunokinin, syringaresinol), and coumarins
(scopole-tin, umbelliferone).[15] Nevertheless, most of the
pre-ceding phytochemical studies on Haplophyllum genus mostly focused on isolation and not on metabolite profiling. By using an ultra-high performance liquid
chromatography (UPLC)-ESI-Q-TOF-MS/MS method,
Hamdi et al.[16] identified 16 compounds in the ethyl
acetate and butanol extracts of H. tuberculatum leaves, of which phenolic acids (vanillic acid, caffeic acid), flavonoids (isorhamnetin, syringetin-3-glucoside), li-gnans (justicidins A and B, kusunokinin arabelline, majidine), and alkaloids (skimmianine, γ-fagarine, dictamnine, tubacetine derivative).
In order to assess the phytochemical profile of the flowers, roots, and stems of H. sahinii and H.
vulcan-icum, a conventional extraction procedure with
ethanol was performed as the first step (Table 1). After that, the qualitative analysis of these extracts was carried out by HPLC-DAD-ESI-Q-TOF-MS/MS. A Q-TOF mass analyzer was used since it provides accurate mass measurements and isotopic fidelity which allows obtaining the molecular formula (MF) of target com-pounds. Consequently, the high-resolution mass data (HR-MS) were in agreement with the theoretical molecular formula (mass error < 5 ppm). Figure 1 depicts the representative base peak chromatogram (BPC) of analyzed Haplophyllum samples, whereas
Table 2 presents the retention times (tR), experimental
m/z of precursor ions, MF, main MS/MS fragments, and
proposed assignments of labeled peaks. This way, a number of 34 phytochemical metabolites were tenta-tively annotated, including flavonoids, furoquinoline and amide alkaloids, phenylpropanoid, arylnaphtha-lene, and dibenzobutyrolactone lignans along with furano- and prenylated coumarins. Two compounds
were further confirmed by comparing their tR, UV, MS,
and MS/MS spectra with those of pure standards; for the rest, their tentative identification was performed by analyzing the UV, HR-MS spectra, and MS/MS fragmentation patterns and comparing them with
Table 1. Spectral data of compounds tentatively identified in the flower, root, and stem extracts of H. sahinii and H. vulcanicum. [a] No. tR [min] UV [nm] [M +H] + exp. [m/z ] [M +H] + calcd. [m/z ] δ [ppm] MF ESI-MS/MS fragments [m/z ] Proposed identity Class Area [%] HSF HSR HSS HVF HVR HVS 1 1.2 260 274.1442 274.1438 1.57 C16 H19 NO 3 256.1365, 202.0819, 172.0754, 144.0828 Unknown – 0.87 1.75 0.80 0.45 1.96 0.83 2 1.4 260 325.2229 325.2221 2.53 C15 H32 O7 233.1646, 163.0368 Unknown – 1.60 1.78 2.57 1.06 1.11 2.10 3 2.0 255, 345 625.1775 625.1763 1.90 C28 H32 O16 479.1149, 317.0620, 302.0349 Isorhamnetin rutino-side Flavonoid 1.09 1.48 1.33 1.15 1.28 1.94 4 2.5 290 331.1552 331.1540 3.63 C19 H22 O5 313.1391, 295.1310, 243.1050 Unknown – 0.66 nd 1.10 0.34 nd 0.36 5 3.6 260 357.1707 357.1697 3.51 C21 H24 O5 297.1321, 163.0290, 153.1194 Unknown – 0.31 0.13 0.20 nd 0.19 nd 6 3.9 260, 320 373.1654 373.1646 2.24 C21 H24 O6 355.1547, 331.1474, 313.1421, 295.1325, 123.1147 Dihydrokusunokinin Lignan 1.00 1.14 2.23 1.44 0.46 1.38 7 4.7 248, 322 260.0927 260.0917 3.73 C14 H13 NO 4 245.0667, 227.0567, 199.0629 Skimmianine [a] Alkaloid 0.25 9.65 0.49 5.68 10.41 6.80 8 5.0 290 297.1496 297.1485 3.64 C19 H20 O3 255.0935, 227.0682, 199.0337 Unknown – 2.54 nd 2.48 1.30 nd 0.98 9 5.3 242, 315 230.0815 230.0812 1.44 C13 H11 NO 3 215.0388, 200.0164 γ -Fagarine Alkaloid nd 10.14 nd nd 11.95 nd 10 5.4 320 362.1968 362.1962 1.66 C20 H27 NO 5 345.1867, 317.1952, 299.1766 Unknown – nd nd nd 1.70 nd 0.55 11 7.1 – 460.2341 460.2330 2.44 C25 H33 NO 7 346.1588, 239.1402, 211.1456 Unknown – 0.91 nd 0.95 nd nd 1.10 12 7.5 248 200.0712 200.0706 2.99 C12 H10 NO 2 185.0539, 156.0393, 129.0491 Dictamnine Alkaloid nd 0.58 nd nd 1.44 nd 13 7.8 230, 261 555.1524 555.1497 4.87 C28 H26 O12 381.0975, 363.0877, 231.0676, 175.0607, 157.0498, 127.0381 Diphyllin acetyl pen-toside I Lignan nd 0.14 nd nd 0.49 nd 14 8.7 288, 325 380.2084 [M +NH 4 ] + 380.2068 4.52 C20 H26 O6 345.1281, 317.1378, 299.1258, 271.1356, 239.1153, 211.1207 Secoisolariciresinol Lignan 11.63 2.93 14.88 12.00 1.25 12.19 15 8.9 326 272.1289 272.1281 2.88 C16 H17 NO 3 216.0642, 201.0362 Unknown – nd nd nd nd 2.01 nd 16 9.5 310 530.2760 530.2748 2.18 C29 H39 NO 8 470.2204, 452.2094, 410.1989, 392.1921, 294.1235, 289.1599 Tubacetine derivative Alkaloid nd 1.01 nd nd 1.09 nd 17 10.0 287 327.1602 327.1591 3.42 C20 H22 O4 245.0806, 193.0492, 178.0298 Unknown – 2.15 nd 2.42 2.43 nd 4.34 18 10.4 230, 261 555.1508 555.1497 1.98 C28 H26 O12 513.1380, 381.0951, 363.0848, 333.0719, 305.0807, 277.0866 Diphyllin acetyl pen-toside II Lignan nd 2.66 nd nd 3.22 nd 19 11.2 230, 265 381.0986 381.0969 4.53 C21 H16 O16 363.0817, 333.0745, 305.0790, 277.0853 Diphyllin Lignan nd 1.24 nd nd 0.29 nd 20 12.1 325 357.1707 357.1697 2.95 C21 H24 O5 297.1448, 255.0987, 241.0858, 175.0375, 163.0356 Rutamarin [a] Coumarin 16.01 0.96 13.70 nd nd nd 21 13.0 288, 320 394.2227 [M +NH 4 ] + 394.2210 0.76 C21 H28 O6 359.1827, 331.1862, 313.1790, 285.1827, 271.1650, 267.1447, 243.1384, 239.1379, 211.1433 Secoisolariciresinol monomethyl ether Lignan 1.84 nd 3.70 1.98 nd 2.67 22 13.9 325 359.1501 359.1489 3.31 C20 H22 O6 341.1395, 323.1249, 309.1140, 151.1071, 123.1140 107.0878 Haplomyrfolol Lignan nd nd 0.38 1.60 nd 1.32 23 14.9 325 297.1489 297.1485 1.28 C19 H20 O3 229.0814, 201.0578, 187.0526, 163.0364, 135.1141 Unknown Coumarin 2.46 0.22 1.49 2.54 0.21 3.07 24 17.1 230, 285 371.1505 371.1489 4.28 C21 H22 O6 353.1371, 335.1257, 321.1090, 303.0994, 151.0764, 135.0423 Kusunokinin Lignan 0.34 4.55 0.83 3.57 9.58 1.78
Table 1. (cont.) No. tR [min] UV [nm] [M +H] + exp. [m/z ] [M +H] + calcd. [m/z ] δ [ppm] MF ESI-MS/MS fragments [m/z ] Proposed identity Class Area [%] HSF HSR HSS HVF HVR HVS 25 17.9 325 297.1489 297.1485 1.28 C19 H20 O3 229.0814, 201.0578, 187.0526, 163.0364, 135.1141 Unknown – 2.13 nd 1.50 nd nd 0.93 26 18.6 260, 320 311.1653 311.1642 3.64 C20 H22 O3 255.0729, 231.0715, 203.0483, 189.0514, 131.0470, 103.0533 Obtusifolin Coumarin nd 2.27 nd 1.72 nd nd 27 18.9 284, 325 357.1712 357.1696 4.44 C20 H20 O6 339.1247, 321.1140, 307.0742, 247.0481, 223.0563, 208.0344, 135.0430 Haplomyrfolin Lignan 0.83 nd 2.11 1.18 1.32 1.01 28 19.1 325 315.1602 315.1591 3.55 C19 H22 O4 297.1385, 241.0464, 229.0791, 187.0340, 163.0350, 153.1239 Unknown – nd nd nd 1.58 nd 1.87 29 19.6 285, 325 527.2646 527.2639 1.25 C30 H38 O8 509.2504, 357.1299, 153.1250, 135.1127, 107.0823 Haplomyrfolin deriva-tive Lignan 0.55 nd 0.72 0.97 nd 0.56 30 20.8 268 297.1487 297.1485 0.60 C19 H20 O3 229.0814, 201.0578, 187.0526 Unknown – 2.03 0.95 1.10 nd 0.96 nd 31 21.2 322 313.1435 313.1434 0.21 C19 H20 O4 231.0527, 203.0625, 135.0425 Unknown 0.47 0.60 1.04 1.75 1.02 1.83 32 21.5 320 535.1989 535.1963 4.94 C30 H30 O9 373.1704, 275.0890, 221.0460, 175.0313, 153.1267, 135.117 Kusunokinin deriva-tive Lignan 0.24 nd nd nd nd nd 33 21.9 250, 320 414.1928 414.1911 4.08 C23 H27 NO 6 380.1834, 246.0743, 231.0535, 216.0261 Dehydrohaplotubine Alkaloid 1.33 0.92 nd 0.55 1.02 2.27 34 22.7 320 351.0870 351.0863 1.96 C20 H14 O6 333.0763, 297.1493, 215.0640 Unknown – 2.76 nd 0.82 nd nd nd 35 22.8 250, 320 414.1928 414.1911 4.08 C23 H27 NO 6 380.1834, 246.0743, 231.0535, 216.0261 Dehydrohaplotubine isomer Alkaloid nd 0.54 nd 0.49 0.45 1.09 36 23.1 325 408.2393 [M +NH 4 ] + 408.2381 3.17 C22 H30 O6 345.1697, 313.1433, 285.1459, 239.1444, 197.1298, 155.0862 Secoisolariciresinol dimethyl ether Lignan 0.97 2.89 1.69 0.97 2.03 0.73 37 23.3 285, 325 492.2972 492.2956 3.30 C27 H41 NO 7 432.2738, 372.2521, 354.2416, 271.1682, 253.1592, 222.1489 Dihydrotubasenicine Alkaloid 0.48 nd nd 1.35 nd 1.42 38 23.7 285, 325 490.2809 490.2799 1.78 C27 H39 NO 7 430.2627, 370.2397, 271.1679, 253.1565, 220.1259 Tubasenecine Alkaloid 2.83 1.66 nd 3.24 1.03 1.47 39 24.0 325 408.2393 [M +NH 4 ] + 408.2381 3.17 C22 H30 O6 391.2117, 373.2048, 345.1814, 317.1772, 299.1682, 285.1489 Secoisolariciresinol dimethyl ether Lignan 6.99 1.30 12.90 5.17 1.29 8.11 40 24.8 320 336.2184 [M +NH 4 ] + 336.2169 4.60 C19 H26 O4 301.2240, 273.2256, 241.1952, 213.1937, 199.1779 Unknown – nd nd 1.30 2.65 0.26 nd 41 25.0 285, 325 512.2662 512.2643 3.76 C29 H37 NO 7 452.2396, 392.2205, 374.2086, 322.1765, 271.1697, 242.1159, Tubacetine Alkaloid 5.18 8.75 3.07 3.39 6.87 3.24 42 26.7 – 798.4091 [M +NH 4 ] + 798.4059 4.08 C43 H56 O13 763.3598, 639.3318, 551.2209, 509.2896, 389.1940, 357.1676 Unknown – 0.26 0.44 0.36 0.23 0.68 0.20 43 27.6 285, 325 476.3006 476.3007 0.14 C27 H41 NO 6 416.2754, 356.2553, 300.1900, 274.1774, 255.1690, 222.1451 Dihydrotubasenecine derivative Alkaloid 0.94 0.77 0.95 0.95 0.48 0.62
Table 1. (cont.) No. tR [min] UV [nm] [M +H] + exp. [m/z ] [M +H] + calcd. [m/z ] δ [ppm] MF ESI-MS/MS fragments [m/z ] Proposed identity Class Area [%] HSF HSR HSS HVF HVR HVS 44 28.0 – 724.3728 [M +NH 4 ] + 724.3691 3.77 C40 H50 O11 689.3360, 672.3141, 611.2997, 509.2845, 477.1876, 449.2654 Secoisolariciresinol dimer I Lignan 1.32 0.42 0.80 1.25 0.18 1.40 45 28.7 320 524.2766 [M +NH 4 ] + 524.2748 3.35 C30 H36 O8 357.1347, 135.1151, 109.1001 Haplomyrfolin deriva-tive Lignan nd 3.51 0.28 0.88 2.17 0.37 46 29.5 320 520.1991 [M +NH 4 ] + 520.1966 4.99 C29 H26 O8 353.1015, 307.0892, 231.0635, 173.0612, 135.0427 Kusunokinin deriva-tive Lignan nd nd 0.18 nd nd 0.41 47 29.8 210, 325 345.1700 345.1696 1.11 C20 H24 O5 209.0450, 194.0195 Geranyloxyhydroxy methoxycoumarin Coumarin 1.37 1.50 0.49 1.26 1.67 1.40 48 30.7 – 588.2824 [M +NH 4 ] + 588.2803 3.64 C31 H38 O10 367.1346, 283.1120, 245.0887, 299.0600, 201.0997, 185.0698 Unknown – nd 1.69 nd nd 1.39 nd 49 30.9 290, 334 329.1752 329.1747 2.33 C20 H24 O4 193.0483, 178.0266 Geranyloxy methoxy-coumarin Coumarin 2.79 nd 0.19 1.32 nd 1.37 50 31.3 – 572.2493 [M +NH 4 ] + 572.2490 0.50 C30 H34 O10 351.0873, 267.0595, 229.0505, 185.0587, 135.0392 Unknown – nd 1.21 0.21 0.93 1.45 nd 51 32.2 – 724.3725 [M +NH 4 ] + 724.3750 4.76 C40 H50 O11 707.3400, 679.3357, 363.1753, 345.1697, 317.1721, 299.1652 Secoisolariciresinol dimer II Lignan 7.03 1.61 6.17 8.09 1.33 6.61 52 33.7 – 738.3866 [M +NH 4 ] + 738.3848 2.52 C41 H52 O11 693.3552, 359.1825, 345.1670, 331.1872, 371.1740, 285.1835 Secoisolariciresinol mixt dimer Lignan 0.85 nd 0.72 0.79 nd 0.50 53 34.1 248, 320 382.2016 382.2013 0.83 C23 H27 NO 4 246.0751, 231.0515, 216.0228 Haplotubine deriva-tive Alkaloid nd 2.90 nd 0.86 2.61 3.58 54 34.4 225, 325 359.1861 359.1853 2.23 C21 H26 O5 223.0581, 208.0341, 193.0114 Geranyloxy dime-thoxycoumarin Coumarin 3.68 2.50 0.38 2.03 1.34 2.68 55 34.8 – 378.2438 378.2428 2.77 C25 H31 NO 2 257.1867, 242.1122, 105.0307 Unknown – 0.41 1.80 nd 0.19 0.56 nd [a] Identification performed by comparison with standard; HSF H. sahinii flower extract; HSR H. sahinii root extract; HSS H. sahinii stem extract; HVF H. vulcanicum flower extract; HSV H. vulcanicum root extract; HVS H. vulcanicum stem extract; MF molecular formula; nd not detected; tR retention time.
available literature or databases (KNApSAck, Chem-Spider, PubChem or Metlin).
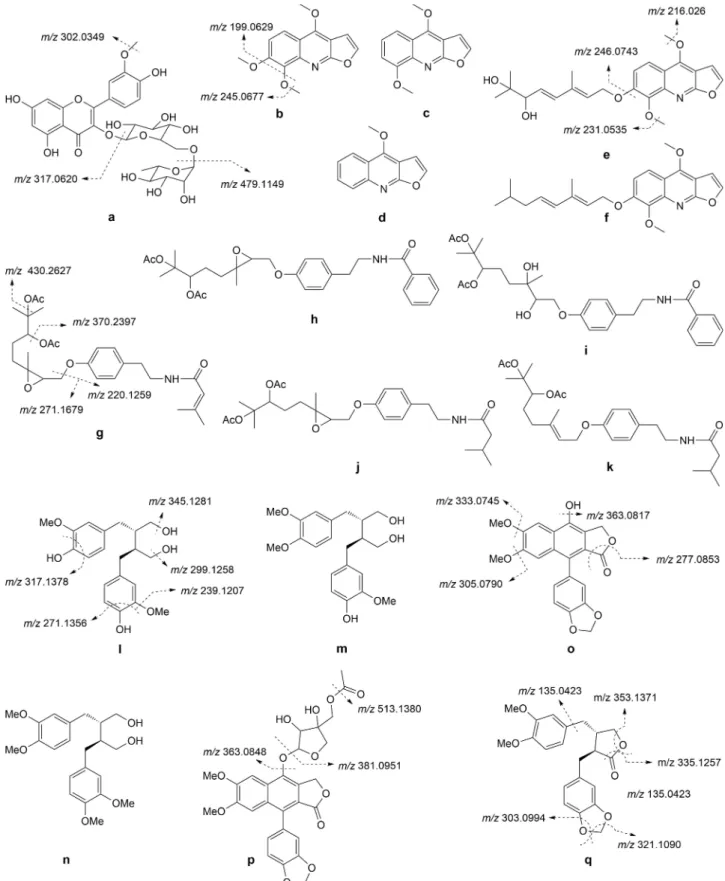
(a) Flavonoids
A single flavonoid was spotted in Haplophyllum
extracts. The precursor ion at m/z 625.1775 (C28H32O16)
exhibited neutral losses of rhamnosyl, glucosyl, and methyl groups, respectively, yielding fragment ions at
m/z 479.1149 ([M rhamnosyl + H]+), 317.0620
([agly-cone + H]+), and 302.0349 ([aglycone CH3+H]+). This
led to the tentative identification of peak 3 as isorhamnetin rutinoside (Figure 2a), a constituent pre-viously reported in various Haplophyllum species, such
as H. pedicellatum, H. robustum, and H. glabrum.[17]
Figure 1. Representative base peak chromatogram of the flower, root, and stem extracts of H. sahinii and H. vulcanicum.
Table 2. Inhibitory activity (inhibition % � S.D.; IC50values) of the ethanol extracts of two Haplophyllum species against AChE,
BChE, and TYR at 200 μg/mL. Species screened Plant
parts
AChE inhibitory activity (Inhibition % � S.D.[a])
BChE inhibitory activity (Inhibition % � S.D.)
TYR inhibitory activity
(Inhibition % � S.D.) Haplophyllum sahinii Root 34.85 � 0.58 53.01 � 2.68
(IC50=190.30 � 5.60) 3.60 � 1.27 Stem 17.90 � 3.84 73.02 � 0.85 (IC50=64.93 � 1.38) 30.68 � 1.19 Flower 16.63 � 3.29 75.54 � 0.38 (IC50=79.67 � 0.52) 34.41 � 0.83 Haplophyllum vulcanicum Root 50.71 � 5.50 51.05 � 5.76
(IC50=200.90 � 0.14) 1.88 � 0.76 Stem 36.05 � 3.96 (IC50=203.18 � 5.33) 79.05 � 4.34 (IC50=87.53 � 2.05) 6.87 � 1.76 Flower 18.68 � 4.45 69.71 � 3.50 (IC50=97.57 � 3.49) 25.19 � 1.62 References[b] 92.14 � 2.49 (IC50=0.68 � 0.05) 81.93 � 2.52 (IC50=34.44 � 0.82) 89.71 � 0.54
Figure 2. Possible chemical structures of compounds tentatively identified in the flower, root, and stem extracts of H. sahinii and H.
(b) Alkaloids
Furoquinoline Alkaloids
Six furoquinoline alkaloids were identified in
Haplo-phyllum extracts. Skimmianine (peak 7, Figure 2b) was
unequivocally assigned by comparing its spectral data with those of a commercially available standard and
literature data.[18] Its MS/MS spectrum revealed the
following fragment ions at m/z 245.0667, 227.0567, and 199.0629 that were produced by the sequential
neutral losses of CH3, H2O, and CO from the precursor
ion [M + H]+ at m/z 260.0927. Compound 9 ([M + H]+
at m/z 230.0815, C13H11NO3) showed two suggestive
MS/MS fragment ions at m/z 215.0388 ([M CH3+H]+)
and 200.0164 ([M 2 CH3+H]+), being tentatively
assigned to γ-fagarine (Figure 2c). As compared to this,
peak 12 ([M + H]+ at m/z 200.0712, C12H10NO2)
indicated the presence of only one methoxy group
(fragment ion at m/z 185.0539 ([M CH3+H]+), which
was in agreement with the structure of dictamnine (Figure 2d). Skimmianine, β-fagarine, and dictamnine are the previously known constituents in Haplophyllum
genus including H. vulcanicum.[15,16]
Additionally, three furoquinoline alkaloid deriva-tives (peaks 33, 35, and 53) were observed in the
more non-polar region of the chromatograms (tR>
20 min). Since they all yielded the same fragment ions at m/z 246.0743, 231.0535, and 216.0261 generated by
the sequential losses of the bulky side chains, the first and second methyl group, respectively, haplopine was proposed as their base structure. However, the precise nature of the radical cannot be deduced from the MS/ MS data; furthermore, since the literature survey did not reveal structures to overlap those suggested by the molecular formula and spectral data, the exact structures of these compounds were not elucidated.
Nevertheless, as compared to haplotubine
(C23H29NO6), a dihydroxygeranyl furoquinoline alkaloid
reported in H. tuberculatum,[6] the molecular formula
of the two isomers 33 and 35 (C23H27NO6) suggested a
higher double bond equivalent (DBE) value that could arise from an additional carbon-carbon double bond in the side chain. A possible structure for one of the isomers is shown in Figure 2e. In the case of peak 53, the same DBE as dehydrohaplotubine was noticed
(C23H27NO4), but the molecule contained two less
oxygen atoms that would indicate a fully deoxygen-ated geranyl side chain (Figure 2f).
Amide Alkaloids
Five amide alkaloids were spotted in the Haplophyllum
extracts. The precursor ion [M + H]+ at m/z 490.2809
of peak 38 suggested a molecular formula of
C27H39NO7. The main fragment ions at m/z 430.2627
of one or two acetic acid (C2H4O2, 60 Da) units, respectively. These data were in agreement with the structure of tubasenecine, previously reported in H.
tuberculatum.[6]Furthermore, the fragment ions at m/z
271.1679 and 220.1259 were obtained by the cleavage of the etheric bridge, reinforcing the proposed structure (Figure 2g). The exact same fragmentation patterns were observed in the case of the other four
amide alkaloids, namely peaks 16 ([M + H]+ at m/z
530.2748), 37 ([M + H]+ at m/z 492.2972), 41 ([M + H]+
at m/z 512.2660), and 43 ([M + H]+ at m/z 476.3006)
with the following suggested MF:
C29H39NO8,C27H41NO7, C29H37NO7, and C27H41NO6,
re-spectively. Of these, the literature survey revealed a previously reported structure only for peak 41, namely
that of tubacetine (Figure 2h).[6]The molecular formula
of peak 16, as well as its MS/MS spectra, indicated a tubacetine derivative, in which the epoxy bridge was hydrated to a diol group (Figure 2i). If in the case of peak 37, the fragment ions at m/z 432.2738
([M C2H4O2+H]+), 372.2521 ([M 2 C2H4O2+H]+),
271.1682, and 222.1489 supported a dihydrogenated derivative of tubasenecine (dihydrotubasenecine,
Fig-ure 2j), that is not the case of compound 43. The loss
of the two acetic acid units was also observed in the
fragment ions at m/z 416.2754 ([M C2H4O2+H]+) and
356.2553 ([M 2 C2H4O2+H]+), suggesting, as in the
previous compounds, two acetyl groups in the etheric side chain. However, the retrieval of the fragment ion at m/z 222.1451 indicated the same amide group as in dihydrotubasenecine; consequently, the replacement of the epoxy bridge with a double bond would be plausible in the structure of this derivative, hypothesis also supported by the fragment ion at m/z 255.1690 (Figure 2k).
(c) Lignans
Phenylpropanoid Lignans
Peak 14 showed an [M + NH4]+ ion at m/z 380.2084
that pointed out a molecular formula of C20H26O6. The
precursor ion yielded the following fragment ions at
m/z 345.1281 ([M H2O + H]+), 317.1378 ([M H2O CO
+H]+), 299.1258 ([M 2 H2O CO + H]+), 271.1356
([M 2 H2O 2 CO + H]+), and 239.1153 ([M 2 H2O 2
CO CH2O + H]+). As suggested by Liu et al.,[19] these
spectral data might indicate the structure of secoiso-lariciresinol (Figure 2l), a phenylpropanoid lignan
pre-viously isolated from H. tuberculatum.[20]Following the
same fragmentation patterns, two methylated secoiso-lariciresinol derivatives (secoisosecoiso-lariciresinol
monometh-yl ether (21), and secoisolariciresinol dimethmonometh-yl ether (36)) were tentatively annotated in the extracts of the two Haplophyllum species (Figure 2m and 2n). These compounds were not reported before in the current genus, but they are known constituents of Araucaria
angustifolia[21,22] and Justicia procumbens,[19]
respec-tively. Additionally, several dimers of secoisolariciresi-nol (44, 51, and 52) were also supported by the extracted mass spectral data. Although not as frequent as their monomers, dimeric lignans are documented in
some members of Rutaceae family.[21]
Arylnaphthalene Lignans
Peak 19 with [M + H]+ at m/z 381.0969 (C21H16O16)
showed several diagnostic fragment ions at m/z
363.0817 ([M H2O + H]+), 333.0745 ([M H2O CH2O +
H]+), 305.0790 ([M H2O CH2O CO + H]+), and
277.0853 ([M H2O CH2O 2 CO + H]+) that led to the
structure of diphyllin (Figure 2o), a constituent widely
distributed in Haplophyllum genus.[6]In comparison to
this, the precursor [M + H]+ ions at m/z 555.1524 and
555.1508 (with the same molecular formula, C28H26O12)
of peaks 13 and 18 initially suffered the sequential neutral losses of acetyl and pentosyl groups; the obtained fragment ion with m/z at 381.0809 next followed the fragmentation patterns described above for diphyllin. Therefore, they were considered as acetyl-glycosylated diphyllin derivatives. One of these structures might possibly belong to mono-O-acetyldi-phyllin apioside (Figure 2p), a lignan previously
re-ported in H. buxbaumii.[6]
Dibenzylbutyrolactone Lignans
Peak 24 with [M + H]+ at m/z 371.1505 (C21H22O6) was
tentatively assigned to kusunokinin (Figure 2q) based on the fact that it showed similar fragment ions as
those reported by Hamdi et al.,[16] namely (m/z):
353.1371 ([M H2O + H]+), 335.1257 ([M 2 H2O + H]+),
321.1090 ([M 2 H2O CH2+H]+), 303.0994
([M 3 H2O CH2+H]+), and 135.0423 ([C8H7O2]+). By
giving similar fragmentation patterns as those
indi-cated for kusunokinin, peaks 6 ([M + H]+ at m/z
373.1646), 22 ([M + H]+) at m/z 359.1501), and 27 ([M
+H]+ at m/z 357.1712) were tentatively labeled as
dihydrokusunokinin (Figure 2r), haplomyrfolol
(Fig-ure 2s), and haplomyrfolin (Fig(Fig-ure 2t), respectively.
Kusunokinin, haplomyrfolin, and haplomyrfolol were all previously reported in H. vulcanicum, as well as in other Haplophyllum species (H. acutifolium or H.
(d) Coumarins
One furanocoumarin (20) and four prenylated coumarins (26, 47, 49, and 54) were characterized in the extracts of the two Haplophyllum species. The structure of com-pound 20 was confirmed as rutamarin (Figure 2u) by comparing the UV, HR-MS, and MS/MS data with those of standard. Consequently, after deacetylation and cleavage of the side chains, the base peak at m/z 357.1707 generated the fragment ions at m/z 297.1448
([M C2H4O2+H]+), 255.0987 ([M C2H4O2 C3H6+H]+),
and 241.0858 ([M C2H4O2 C4H8+H]+). Peak 26 ([M +
H]+ at m/z 311.1653) was tentatively assigned to
obtusifolin based on its fragment ions with m/z at
255.0729 ([M C4H8+H]+), 231.0715 ([M C5H6 CH2+
H]+), and 203.0483 ([M C5H6 CH2 CO + H]+)
(Fig-ure 2v). Three more coumarin derivatives were observed
and they were considered as geranyloxy derivatives of hydroxymethoxycoumarin (47), methoxycoumarin (49), and dimethoxycoumarin (54). The losses of geranyl
(C10H16, 136 Da) and methyl (CH3, 15 Da) groups were
the diagnostic cleavages in all of these compounds (Figure 2w 2y). Of all these coumarins, only obtusifolin
(26) was previously reported in H. obtusifolium.[3]
Never-theless, furano- and prenylated coumarins are the known constituents of Haplophyllum genus with representatives, such as columbianetin, angelicin, umbelliprenin, 7-iso-prenyloxycoumarin, and osthenol in H. patavinum,
ptilostol, ptilostin, and ptilin in H. ptilostylum.[6]
The metabolite profile of the two Haplophyllum species investigated in the current H. sahinii and H.
vulcanicum was characteristic to the genus pointing
out to presence of alkaloids, lignans, and coumarins as phytochemical markers. Generally, no significant qual-itative differences were observed interspecies. Perhaps the most outstanding remark is the fact that rutamarin (20) was detected only in H. sahinii, a recently identified endemic species. However, more important features can be withdrawn in regard to the botanical part. For instance, the roots concentrated a higher number of alkaloids with γ-fagarine, dictamnine, and several haplotubine derivatives being found only in the root extracts. On the contrary, lignans and coumarins were found to be slightly more abundant in the flower and stem extracts.
Enzyme Inhibitory Activity of the Extracts
In the present study, the inhibitory effects of the extracts obtained from the root, stem, and flowers of two Haplophyllum species, which were screened against AChE, BChE, and TYR at 200 μg/mL, are
displayed in Table 2. Among the extracts screened, the
stem extract of H. sahinii (50.71 � 5.50 %; IC50=
203.18 � 5.33 μg/mL), exhibited the highest inhibition against AChE. Considering both of the species, the
flower extract of H. sahinii (75.54 � 0.38 %; IC50=
79.67 � 0.52 μg/mL) and the stem extract of H.
vulcan-icum (79.05 � 4.34 %; IC50=87.53 � 2.05 μg/mL) were
the most effective extracts in BChE inhibition. The flower and stem extracts of H. sahinii (34.41 � 0.83 % and 30.68 � 1.19 %) inhibited TYR at the highest rate, which can be though considered as low to moderate as found below 50 %.
According to our literature survey, only one single study on ChE and TYR inhibitory activity on
Haplophyl-lum species as one the most medicinally and
econom-ically important genera of Rutaceae family has been
reported so far.[23] In that study, the petroleum ether,
ethyl acetate, methanol, and water extracts from H.
myrtifolium growing in Turkey were tested and the
petroleum ether extract was found to be with the most effective inhibition as compared to that of galantamine as the reference drug. On the other hand, the ethyl acetate extract of this plant was shown to possess a moderate level of TYR inhibition (48.79 mg kojic acid equivalent/g extract) which was observed to have roughly 4–6 folds higher inhibition than those of the water and methanol extracts. In this regard, low to moderate TYR inhibitory effect of two Haplophyllum species screened could be linked to their phenolic
content as proven immensely by numerous studies.[24,25]
Among plant secondary metabolites, alkaloids are proven to possess a marked level of anticholinesterase
effect.[26] For instance, the alkaloids, skimmianine,
γ-fagarine, and dictamnine isolated from the roots of
Zanthoxylum nitidum and also found in Haplophyllum
species, only skimmianine (91.3 � 2.0 %, IC50=8.6 �
0.7 μg/mL) showed a strong AChE inhibitory activity at concentration of 100 μg/mL as compared to the
reference drug; physostigmine (IC50=0.013 �
0.002 μg/mL),[27]which was later confirmed in another
study for skimmianine (58.63 � 1.22 %, IC50=8.52 �
0.64 μg/mL).[28] On the other hand, skimmianine
obtained from the leaves of Evodia lepta was reported to demonstrate a weak level of AChE and BChE
inhibition (IC50=69.1 μM and 5.6 μM against AChE and
IC50=130.2 μM and 26.3 μMagainst BChE for
skimmia-nine and galantamine, respectively).[29] The moderate
inhibition of AChE by skimmianine (67.213 � 0.298 %) as well as dictamnine plus γ-fagarine (52.002 �
0.203 %) was also shown by Cabral et al.,[30] as well as
Cardoso-Lopez et al.[31] Since presence of these
H. vulcanicum and H. sahinii screened herein,
skimmia-nine in particular could be considered to contribute ChE inhibitory effect of these species.
On the other hand, coumarins are known to possess a selective inhibition towards BChE as verified
by many researchers.[32–34] Consistently, the genus
Haplophyllum has been reported to contain a number
of furano- and prenylated coumarins, i. e., columbiane-tin, angelicin, umbelliprenin,
7-isopentenyloxycoumar-in, osthenol, ptilostol, ptilost7-isopentenyloxycoumar-in, etc.[6] Among them,
umbelliprenin was explicated with a strong AChE
inhibitory effect,[35] while we identified various other
selective BChE-inhibiting coumarin derivatives, e. g., umbelliferone, 4-methylumbelliferone, 4-hydroxycou-marin, scopoletin, imperatorin, isoimperatorin,
xantho-toxin, bergapten, and pteryxin.[36 –38] Therefore,
ChE-inhibiting property of H. vulcanicum and H. sahinii could be also possibly related to their coumarin content. However, rutamarin as a furanocoumarin derivative detected in H. sahinii was reported to be
ineffective against AChE with an IC50value of > 500 μM
in a previous study.[39]Thus, this compound seems not
to be associated with AChE inhibition by the inves-tigated Haplophyllum species.
Conclusions
Although the genus Haplophyllum contains medici-nally important species, only one species of this genus has so far been investigated for its ChE and TYR inhibitory potential. In the current study, ChE and TYR inhibitory capacities of the ethanol extracts of H.
sahinii, a new species endemic to Turkey as well as H. vulcanicum have been reported for the first time. Our
results revealed that these two species are the impending sources of particularly ChE-inhibiting mole-cules. Besides, the metabolite profiles of these two species have been elucidated for the first time herein using a highly hyphenated technique. Therefore, this is the first report on in vitro ChE and TYR enzyme inhibitory activities and extensive metabolite profiling of H. vulcanicum and H. sahinii.
Experimental Section
Reagents
LC grade acetonitrile, formic acid, and water were purchased from J. T. Baker (Deventer, The Nether-lands). Skimmianine and rutamarin were provided by PhytoLab (Vestenbergsgreuth, Germany). Electric eel
AChE (type-VI-S, EC 3.1.1.7), horse serum BChE (EC 3.1.1.8), TYR (EC 1.14.1.8.1, 30 U, mushroom tyrosi-nase), 5,5’-dithiobis(2-nitrobenzoic) acid (DTNB),
ace-tylthiocholine iodide, butyrylthiocholine chloride, L
-DOPA, galantamine, and α-kojic acid were acquired from Sigma Aldrich (St. Louis, MO, USA).
Plant Materials
The samples of Haplophyllum sahinii and H. vulcanicum were collected from different areas of Konya Province in Turkey, in May 2017. The plants were identified by Prof. Dr. Osman Tugay from the Department of Pharmaceutical Botany, Faculty of Pharmacy, Selcuk University (Konya, Turkey). The voucher specimens are deposited with the KNYA Herbarium of the Depart-ment of Biology, Faculty of Sciences, Selçuk University (Konya, Turkey) with the following herbarium num-bers, e. g., KNYA Herb. No. 28.388 and KNYA Herb. No. 28.389, respectively.
Extraction Procedure
The fresh plant materials were firstly separated into the aerial and root parts, dried in shade at room temperature and ground to a fine powder in a mechanic grinder. Each powdered plant part (e. g., flowers, roots, and stems; accurately weighed between 20 –5 g) was separately extracted with ethanol (80 %) and filtered. After filtration, the organic phases were evaporated using a rotary evaporator (Büchi, Flawil,
Switzerland) at 45°C to dryness. The ethanol extracts
were obtained and kept in the freeze until the experiments were performed.
HPLC-DAD-ESI-Q-TOF-MS/MS Analysis
An Agilent 1200 Series HPLC system (Agilent Technol-ogies, Palo Alto, CA, USA) equipped with a G1329B auto-sampler, G1312B binary pump, G1315B diode array detector (DAD), and 6510 A electrospray ioniza-tion-quadrupole-time-of-flight-mass spectrometer (ESI-Q-TOF-MS) was employed for the HPLC-DAD-ESI-Q-TOF-MS/MS analysis. The chromatographic separation
was performed on a Phenomenex Gemini C18 (100 ×
mm i.d., 3 μm) column, with a stepwise gradient composed of 0.1 % formic acid in water (A) and 0.1 % formic acid in acetonitrile (B); 0–10 min–40 % B, 10– 30 min– 40–80 % B, 30– 35 min–80 % –90 % B; flow-rate 0.2 mL/min. The following ESI-Q-TOF-MS parame-ters were used in the positive ion mode: mass range
12 L/min; nebulizer pressure 40 psi; skimmer 65 V; capillary voltage 4000 V; fragmentor 120 V; collision-induced dissociation energies 10 and 40 V.
Microtiter Assay for AChE and BChE Inhibition
AChE and BChE inhibitory activities of the extracts were determined by the slightly modified spectrophotometric
method developed by Ellman et al.[40] Briefly, 14 μL of
sodium phosphate buffer (pH 8.0), 1 μL of DTNB, 2 μL of test solution, and 2 μL of AChE/BChE solution were added by multichannel automatic pipette (Gilson Pipet-man, France) in a 96-well microplate and incubated for
10 min at 25°C. The reaction was then initiated with the
addition of 1 μL of acetylthiocholine iodide/butyrylthio-choline chloride. Hydrolysis of acetylthioiodide/butyrylthio-choline iodide/ butyrylthiocholine chloride was monitored by the for-mation of the yellow 5-thio-2-nitrobenzoate anion as a result of the reaction of DTNB with thiocholine, catalyzed by enzymes at 412 nm utilizing a 96-well microplate reader (VersaMax Molecular Devices, USA). The measure-ments and calculations were evaluated by using Softmax PRO 4.3.2.LS software. The percentage of inhibition of AChE/BChE was determined by comparison of the reaction rates of samples vs. blank sample (ethanol in phosphate buffer, pH 8.0) using the formula I% =
100 [(A1/A2) × 100], where A1 is the absorbance of the
sample solutions at 412 nm and A2 is the average
absorbance of the control solutions at the same wave-length. The results are given as mean � standard devia-tion of the % inhibidevia-tions obtained from three experi-ments. Galantamine was used as the reference drug.
Microtiter Assay for TYR Inhibition
Inhibition of TYR was determined using the modified
dopachrome method withL-DOPA as substrate.[41]The
assay was conducted in a 96-well microplate and an ELISA microplate reader (VersaMax Molecular Devices, USA) was used to measure absorbance at 492 nm. An aliquot of the extracts dissolved in DMSO with 8 μL of
phosphate buffer (pH 6.8), 3 μL of TYR, and 3 μL of L
-DOPA were put in each well. The results were compared with control (DMSO). The percentage of TYR
inhibition was calculated as follows: I% = 100 [(A1/
A2) × 100], where A1 is the absorbance of the sample
solutions at 492 nm and A2is the average absorbance
of the control solutions at the same wavelength. The results are given as mean � standard deviation of the % inhibitions obtained from three parallel experi-ments. α-Kojic acid was used as the reference.
Author Contribution Statement
I. E. O. performed the experiments with enzyme inhibition, analyzed obtained data, and wrote the article. F. S. S. D. performed the experiments with enzyme inhibition. O. T. collected and identified the plant samples. E. K. prepared the extracts and assisted the enzyme inhibition assays. S.V. L. and K. S.-W. performed the metabolic profiling experiments on the extracts and wrote the article. M. S. analyzed the obtained data and assisted with writing the article.
References
[1] O. Tugay, D. Ulukuş, ‘Haplophyllum sahinii (Rutaceae), a new species from Central Anatolia (Turkey)’, Phytotaxa
2017, 297, 265–272.
[2] D. Ulukuş, O. Tugay, ‘Haplophyllum ermenekense (Rutaceae), a new species from Turkey’, Phytokeys 2018, 111, 119 –131. [3] M. Tekin, N. Eruygur, ‘The structural studies on the medicinal plant Haplophyllum telephioides’, Rev. Bras. Farm-acogn. 2016, 26, 544–552.
[4] I. Bessonova, D. Kurbanov, S. Y. Yunusov, ‘Components of Haplophyllum ramosissimum’, Chem. Nat. Compd. 1989, 25, 39 –40.
[5] S. Hadjadj, Z. Bayoussef, A. O. El Hadj-Khelil, H. Beggat, Z. Bouhafs, Y. Boukaka, I. A. Khaldi, S. Mimouni, F. Sayah, M. Tey, ‘Ethnobotanical study and phytochemical screening of six medicinal plants used in traditional medicine in the Northeastern Sahara of Algeria (area of Ouargla)’, J. Med. Plants Res. 2015, 9, 1049 –1059.
[6] A. E. Al-Snafi, ‘Pharmacological importance of Haplophyl-lum species grown in Iraq – a review’, IOSR J. Pharm. 2018, 8, 54 –62.
[7] A. Ulubelen, M. Ozturk, ‘Alkaloids, coumarins and lignans from Haplophyllum species’, Rec. Nat. Prod. 2008, 2, 54 –69. [8] G. Orhan, I. Orhan, N. Subutay-Oztekin, F. Ak, B. Sener, ‘Contemporary anticholinesterase pharmaceuticals of natu-ral origin and their synthetic analogs for the treatment of Alzheimer’s disease’, Recent Pat. CNS Drug Discovery 2009, 4, 43 –51.
[9] O. Ustun, F. S. Senol, M. Kurkcuoglu, I. E. Orhan, M. Kartal, K. H. C. Baser, ‘Investigation on chemical composition, anticholinesterase and antioxidant activities of extracts and essential oils of Turkish Pinus species and pycnogenol’, Ind. Crops Prod. 2012, 38, 115 –123.
[10] L. S. Schneider, ‘Treatment of Alzheimer’s disease with cholinesterase inhibitors’, Clin. Geriatr. Med. 2001, 17, 337 – 358.
[11] A. Alias, M. Al-Zubaidy, Y. Mousa, F. Mohammad, ‘Plasma and whole brain cholinesterase activities in three wild bird species in Mosul, IRAQ: In Vitro inhibition by insecticides’, Interdiscip. Top. Gerontol. 2011, 4, 144 –148.
[12] L. Maggi, R. Mantegazza, ‘Treatment of myasthenia gravis’, Clin. Drug Invest. 2011, 31, 691 –701.
[13] I. E. Orhan, G. Orhan, E. Gurkas, ‘An overview on natural cholinesterase inhibitors - a multi-targeted drug class - and
their mass production’, Mini-Rev. Med. Chem. 2011, 11, 836 –842.
[14] E. Mendes, M. de J. Perry, A. P. Francisco, ‘Design and discovery of mushroom tyrosinase inhibitors and their therapeutic applications’, Exp. Opin. Drug Discov. 2014, 9, 533 –554.
[15] I. E. Orhan, M. T. H. Khan, ‘Flavonoid derivatives as potent tyrosinase inhibitors – a survey of recent findings between 2008 –2013’, Curr. Top. Med. Chem. 2014, 14, 1486 –1493. [16] B. Gözler, D. Rentsch, T. Gözler, N. Ünver, M. Hesse,
‘Lignans, alkaloids and coumarins from Haplophyllum vulcanicum’, Phytochemistry 1996, 42, 695–699.
[17] A. Hamdi, J. Viane, M. A. Mahjoub, K. Majouli, M. H. H. Gad, M. Kharbach, K. Demeyer, Z. Marzouk, Y. V. Heyden, ‘Polyphenolic contents, antioxidant activities and UPLC-ESI-MS analysis of Haplophyllum tuberculatum A.Juss leaves extracts’, Int. J. Biol. Macromol. 2018, 106, 1071 –1079. [18] A. Ulubelen, S. Oksuz, B. Halfon, Y. Aynehchi, T. J. Mabry,
S. A. Matlin, ‘Flavonoids from Haplophyllum pedicellatum, H. robustum and H. glabrinum’, Phytochemistry 1984, 12, 2941 –2943.
[19] E. Kozioł, S. V. Luca, L. Marcourt, M. Nour, E. Hnawia, J. Jakubowicz-Gil, R. Paduch, T. Mroczek, J. L. Wolfender, K. Skalicka-Woźniak, ‘Efficient extraction and isolation of skimmianine from New Caledonian plant Medicosma leratii and evaluation of its effects on apoptosis, necrosis, and autophagy’, Phytochem. Lett. 2019, 30, 224 –230.
[20] B. Liu, Y. Yang, H. Liu, Z. Xie, Q. Li, M. Deng, F. Li, J. Peng, H. Wu, ‘Screening for cytotoxic chemical constituents from Justicia procumbens by HPLC-DAD-ESI-MS and NMR’, Chem. Cent. J. 2018, 12, article No. 6.
[21] D. T. A. Youssef, ‘Lignans from the aerial parts of Haplophyl-lum tuberculatum (Forssk) A. Juss.’, Bull. Pharm. Sci. Assiut Univ. 2005, 28, 261–267.
[22] S. F. Fonseca, L. T. Nielsen, E. A. Ruveda, ‘Lignans of Araucaria angustifolia and 13C-NMR analysis of some
phenyltetralin lignans’, Phytochemistry 1979, 18, 1703 – 1708.
[23] X.-J. Zhou, X.-L. Chen, X.-S. Li, J. Su, J.-B. He, Y.-H. Wang, Y. Li, Y.-X. Cheng, ‘Two dimeric lignans with an unusual α,β-unsaturated ketone motif from Zanthoxylum podocarpum and their inhibitory effects on nitric oxide production’, Bioorg. Med. Chem. Lett. 2011, 21, 373–376.
[24] G. Zengin, C. Sarıkurkcu, A. Aktumsek, R. Ceylan, O. Ceylan, ‘A comprehensive study on phytochemical characterization of Haplophylum myrtifolium Boiss. endemic to Turkey and its inhibitory potential against key enzymes involved in Alzheimer, skin diseases and type II diabetes’, Ind. Crops Prod. 2014, 53, 244–251.
[25] M. T. H. Khan, ‘Novel tyrosinase inhibitors from natural resources – their computational studies’, Curr. Med. Chem.
2012, 19, 2262 –2272.
[26] S. Y. Lee, N. Baek, T.-G. Nam, ‘Natural, semisynthetic and synthetic tyrosinase inhibitors’, J. Enzyme Inhib. Med. Chem.
2016, 31, 1 –13.
[27] K. Habartová, L. Cahlíková, M. Řezáčová, R. Havelek, ‘The biological activity of alkaloids from the Amaryllidaceae: from cholinesterases inhibition to anticancer activity’, Nat. Prod. Commun. 2016, 11, 1587 –1594.
[28] Z.-D. Yang, D.-B. Zhang, J. Ren, M.-J. Yang, ‘Skimmianine, a furoquinoline alkaloid from Zanthoxylum nitidum as a
potential acetylcholinesterase inhibitor’, Med. Chem. Res.
2012, 21, 722 –725.
[29] J. Sichaem, A. Jirasirichote, K. Sapasuntikul, S. Khumkratok, P. Sawasdee, T. M. L. Do, S. Tip-Pyang, ‘New furoquinoline alkaloids from the leaves of Evodia lepta’, Fitoterapia 2014, 92, 270 –273.
[30] R. S. Cabral, M. C. Sartori, I. Cordeiro, C. L. Queiroga, M. N. Eberlin, J. H. Lago, P. R. Moreno, M. Young, ‘Anticholinester-ase activity evaluation of alkaloids and coumarin from stems of Conchocarpus fontanesianus’, Rev. Bras. Farm-acogn. 2012, 22, 374–380.
[31] E. M. Cardoso-Lopes, J. A. Maier, M. R. da Silva, L. O. Regasini, S. Y. Simote, N. P. Lopes, J. R. Pirani, V. da S. Bolzani, M. C. M. Young, ‘Alkaloids from stems of Esenbeck-ia leiocarpa Engl. (Rutaceae) as potentEsenbeck-ial treatment for Alzheimer disease’, Molecules 2010, 15, 9205 –9213. [32] I. E. Orhan, H. O. Gulcan, ‘Coumarins: auspicious
cholines-terase and monoamine oxidase inhibitors’, Curr. Top. Med. Chem. 2015, 15, 1673 –1682.
[33] L. G. de Souza, M. N. Rennã, J. D. Figueroa-Villar, ‘Coumar-ins as cholinesterase inhibitors: A review’, Chem.-Biol. Interact. 2016, 254, 11– 23.
[34] S. Karakaya, M. Koca, S. V. Yilmaz, K. Yildirim, N. M. Pinar, B. Demirci, M. Brestic, O. Sytar, ‘Molecular docking studies of coumarins isolated from extracts and essential oils of Zosima absinthifolia Link as potential inhibitors for Alz-heimer’s disease’, Molecules 2019, 24, pii: E722.
[35] S. Dall’Acqua, F. Maggi, P. Minesso, M. Salvagno, F. Papa, S. Vittori, G. Innocenti, ‘Identification of non-alkaloid acetyl-cholinesterase inhibitors from Ferulago campestris (Besser) Grecescu (Apiaceae)’, Fitoterapia 2010, 81, 1208 –1212. [36] I. Orhan, F. Tosun, B. Sener, ‘Coumarin, anthroquinone and
stilbene derivatives with anticholinesterase activity’, Z. Naturforsch. 2008, 63, 366– 370.
[37] F. S. Senol, K. Skalicka-Wozniak, M. T. H. Khan, I. E. Orhan, B. Sener, K. Glowniak, ‘An in vitro and in silico approach to cholinesterase inhibitory and antioxidant effects of the methanol extract, furanocoumarin fraction, and major coumarins of Angelica officinalis L. fruits’, Phytochem. Lett.
2011, 4, 462 –467.
[38] I. E. Orhan, F. S. Senol, S. Shekfeh, K. Skalicka-Wozniak, E. Banoglu, ‘Pteryxin – A promising butyrylcholinesterase-inhibiting coumarin derivative from Mutellina purpurea’, Food Chem. Toxicol. 2017, 109, 970–974.
[39] J. M. Rollinger, D. Schuster, B. Danzl, S. Schwaiger, P. Markt, M. Schmidtke, J. Gertsch, S. Raduner, G. Wolber, T. Langer, H. Stuppner, ‘In silico target fishing for rationalized ligand discovery exemplified on constituents of Ruta graveolens’, Planta Med. 2009, 75, 195–204.
[40] G. L. Ellman, K. D. Courtney, V. Andres, R. M. Featherstone, ‘A new and rapid colorimetric determination of acetylcho-linesterase activity’, Biochem. Pharmacol. 1961, 7, 88– 95. [41] S.-H. Lee, S. Sandesh, S. Shruti, S.-Y. Seo, ‘Potent
antielas-tase and antityrosinase activities of Astilbe chinensis’, Am. J. Pharmacol. Toxicol. 2009, 4, 127– 129.
Received June 15, 2019 Accepted July 12, 2019