T.C
CLE ÜN VERS TES
FEN B
MLER ENST TÜSÜ
MISIR PÜSKÜLÜNDEN (Stylus maydis) POL FENOL OKS DAZ ENZ
N
SAFLA TIRILMASI VE KARAKTER ZASYONU
Nurten ASLAN
YÜKSEK L SANS TEZ
YOLOJ ANAB
M DALI
YARBAKIR
Haziran-2015
I
TE EKKÜR
Yüksek lisans çal malar n her a amas nda beni destekleyen ve her türlü yard mlar esirgemeyen bilimsel azmini örnek ald m çok k ymetli hocam Say n Doç. Dr.Reyhan GÜL GÜVEN’e en derin minnet ve ükranlar sunar m.
Tez çal mam esnas nda her zaman ilgisini ve yard esirgemeyen Say n Prof.Dr. Kemal GÜVEN, Ar . Gör. Dr. Fatma MATPAN BEKLER ve Dr.Ömer ACER’e çok te ekkür ederim. Ayr ca yard mlar ve arkada klar esirgemeyen sevgili arkada lar m Canan GÜLER, Suna VAROL, Hicran YAPAN, Semra T Z, Ali Evren ARSLANO LU, A.Gafur KARACA, Mehmet Nur KÜÇÜK ve HÜSEY N KARABIYIK ve bu tezin en ba ndan beri olmas nda çok büyük katk olan sab rlar ve sevgilerini asla esirgemeyen sevgili aileme sonsuz te ekkürü bir borç bilirim.
II Sayfa TE EKKÜR ………... NDEK LER ……….... ÖZET………... ABSTRACT ………... ZELGE L STES ………... EK L L STES ………... KISALTMALAR VE S MGELER ………... 1. ………... 1.1. Enzimler…..………...……...
1.1.2. Enzim Aktivitesini Etkileyen Faktörler……….……….. 1.1.3. Enzimlerin Kod Numaralar na Göre S fland lmas ………..…..
1.1.4. Oksidoredüktazlar……… 1.2. Polifenol Oksidaz………....……… 1.2.1. Polifenol Oksidaz n Önemi………..….………..
1.2.2. Polifenol Oksidaz n Tabiattaki Da ve Önemi ………...
1.2.3 Polifenol Oksidaz n Substratlar ………....………... 1.3. Esmerle me Reaksiyonlar ………..
1.3.1. Kimyasal Esmerle me Reaksiyonlar ………....……….
1.3.2. Enzimatik Esmerle me Reaksiyonlar ………..……….
1.4. M r……….………..
1.5. M r Püskülü………...………..………
1.6. Enzimlerin Safla rma A amalar ………...………
1.6.1. Hücre Parçalanmas …………...……… 1.6.2. Amonyum Sülfat Çöktürme……….……… 1.6.3. Diyaliz……….………...…. 1.6.4. Ultrafiltrasyon..………..……… ……….….….. 1.6.5. Jel Filtrasyon Kromatografisi…..………... 1.6.6. Elektroforez………..……….…...… 1.7. nhibisyon.………...……… 1.8. Polifenol Oksidaz le Yap lan Çal malar n Özeti………...
I II IV V VI VII VIII 1 1 1 3 4 4 6 6 7 9 9 9 9 10 11 11 12 13 14 15 15 16 16
III
2. MATERYAL VE METOT ………..
2.1. Materyal………..………... 2.1.1. Çal mada Kullan lan Bitki Örneklerinin Temini ve Analize Haz rlanmas ………..
2.1.2. Çal mada Kullan lan Kimyasal Maddeler………..
2.1.3. Çal mada Kullan lan Cihazlar……….... 2.2. Ham Enzim Özütünün Haz rlanmas ………….………...………...
2.3. Polifenol Oksidaz n Safla lmas …..………...……… 2.4. Ultrafiltrasyon………..……… 2.5. Jel Filtrasyon Kromotografisi………..
2.6. Nativ PAGE ile Elektroforetik Analiz……….…………....
2.7. Enzimin Moleküler A rl n Hesaplanmas …..………... 2.8. Spektofotometrik Ölçümler………..………...
2.9. Enzim Aktivitesine pH’ n Etkisi……….…...
2.10. Enzim Aktivitesine S cakl n Etkisi………...… 2.11. Termal naktivasyon………..……….….
2.12. nhibitörlerin Etkileri………... 2.13. Karbonhidratlar n Etkileri...……….… 2.14. Protein çeri i………..…...…. 2.14.1. Kalitatif Protein çeri i……….………... 2.14.2. Lowry Metodu ………...…………..…... 2.15. Polifenol Oksidaz Enzimine Substrat Konsantrasyonunun Etkisi………...………
3. ARA TIRMA SONUÇLARI VE BULGULAR………
3.1. Enzimin Safla lmas ………
3.2. Substrat Spesifikli i………...…. 3.3. Optimum pH………...…... 3.4. Optimum S cakl k……….……....…...
3.5. Termal naktivasyon………..…………..… 3.6. Polifenol Oksidaz Aktivasyonu………..………...….
3.7. Polifenol Oksidaz nhibisyonu………..…... 3.8. Polifenol Oksidaz n Homojenitesi ve Molekül A rl ……….………...
4. SONUÇ VE TARTI MA………...
5. KAYNAKLAR……….……… 6. ÖZGEÇM ……….……….…. 21 21 21 21 21 21 22 22 22 22 23 23 23 24 24 24 24 24 24 25 25 27 27 28 30 32 33 34 35 36 37 39 45
IV
MISIR PÜSKÜLÜNDEN (Stylus maydis) POL FENOL OKS DAZ ENZ
N
SAFLA TIRILMASI VE KARAKTER ZASYONU
Yüksek Lisans TeziNurten ASLAN
Dicle Üniversitesi Fen Bilimleri Enstitüsü Biyoloji Anabilim Dal
2015
Bu çal mada, m r püskülü (Stylus maydis) polifenol oksidaz (PFO) enziminin safla lmas yap lm ve baz kinetik özellikleri incelenmi tir. M r püskülünden ekstrakte edilen polifenol oksidaz, (NH ) SO (amonyum sülfat) çöktürmesi, diyaliz, ultrafiltrasyon ve jel kromatografisi prosodürleri kullan larak safla ld . Safla lan enzim tek bir bant olarak SDS-PAGE’de elde edilmi tir. M r püskülünün PFO enziminin molekül a rl ise yakla k 158kDa civar nda gözlemlenmi tir.
Katekol ve 4-metil katekol substratlar kullan larak, PFO enziminin optimum pH ve cakl k de erleri belirlenmi tir. Polifenol oksidaz aktivitesi 4-metilkatekol ve katekol için 420 nm'de spektofotomtrik olarak ölçülmü tür. Optimum s cakl k katekol için 40°C ve 4-metilkatekol için 30°C bulunmu tur. Optimum pH katekol için 8 ve 4-4-metilkatekol için 6 olarak tespit edilmi tir.
Ayr ca PFO'n n substrat spesifikli i ve inaktivasyonu incelenmi tir. PFO enziminin optimum pH ve s cakl kta katekol ve 4-metil katekol substratlar için Km ve Vmax de erleri Linewear-Burk yöntemi ile bulunmu tur. Vmax/ Km de erlerine göre PFO enzimi için en uygun substrat n katekol oldu u bulunmu tur. Ayr ca katekol kullan ld nda artan inaktivasyon süresine paralel olarak PFO aktivitesinin bariz bir ekilde azald gözlemlenmi tir.
EDTA, SDS ve sodyum azidin PFO aktivitesi üzerine inhibisyon etkisi katekol substrat kullan larak etkisi incelendi. Bu çal mada s ras yla sodyum azidin % 26.6 EDTA’n n % 22.2 ve SDS’nin % 12.2 oran nda enzimi inhibe etti i tespit edildi. Ayr ca sukroz, fruktoz, riboz ve glikoz gibi karbonhidratlar n PFO aktivitesi üzerine etkisi ara lm r. Fruktoz ve ribozun enzimi % 17 oran nda aktive etti i, glikozun % 16, sukrozun ise % 4 oran nda aktive etti i tespit edilmi tir.
V
ABSTRACT
PURIFICATION AND CHARACTERIZATION OF
POLYPHENOL OXIDASE FROM CORN TASSEL(Stylus maydis)
MASTER. ThesisNURTEN ASLAN
Dicle University, Institute of Science, Department of Biology 2015
In this work, purification of polifenol oxidase enzyme from stylus maydis was done and some kinetic characteristics were analysed. Polifenol oxidase extracted from stylus maydis was purified by the procedures of ammonium sulphate (NH ) SO precipitation, dialysis, ultrafiltration and jel chromatography. One band for purified enzyme was obtained by SDS-PAGE. Molecular weight of stylus maydis PPO enzyme was calculated as approximately 158 kDa.
Optimum pH and temperature values of PPO enzyme were determined by using catechol and methyl catechol substrates. Polifenol oxidase activity was measured for 4-methyl catechol ve catechol at 420 nm as spectrophotometric. Optimum temperature was founded as 40°Cfor catechol and 30°C for 4-methyl catechol. Optimum pH was determined as 8 for catechol and 6 for 4-methyl catechol.
Besides, substrat specifity and temperature inactivation of PPO were analysed. PPO enzyme’s Km and Vmax values for catechol and 4-methyl catechol substrates at optimum pH and temperature were determined by using Linewear-Burk method. According to Vmax and Km values, the most suitable substrate was found to be catechol. Furthermore, it was observed that PPO activity decreased clearly in paralel with increasing inactivation time when catechol used.
Inhibition effect of EDTA, SDS and sodium azide on PPO activity was analysed by using catechol substrate. In this work, sodium azide, EDTA and SDS were found to inhibit the enzyme activity as 26.6 %, 22.2 % and 12.2 %, respectively. Besides, the effects of carbohydrates such as sucrose, fructose, ribose and glucose on PPO activity were investigated. The enzyme was found to be activated 17 % by fructose and ribose, 16 % by glucose and 4 % by sucrose.
VI
Çizelge No Sayfa Çizelge 1.1. Enzimlerin kod numaralar na göre s ralanmas
Çizelge 1.2. Polifenol oksidaz enziminin katalizledi i reaksiyonlar Çizelge 1.3. Polifenol oksidaz enziminin baz substratlar n yap
Çizelge 1.4. Fenolik bile ikler Çizelge 3.1. Polifenol oksidaz enziminin Sephadex G-75 jel geçirgenlik
kromatografisi ile elde edilen fraksiyonlar
Çizelge 3.2. Polifenol oksidaz aktivitesi için çe itli substratlarla elde edilmi veriler Çizelge 3.3. Katekol substrat na ait Lineweaver-Burk grafi i
Çizelge 3.4. 4-metilkatekol substrat na ait Lineweaver-Burk grafi i
Çizelge 3.5. M r püskülünden elde edilen polifenol oksidaz aktivitesinin katekol substrat nda pH ile de imi
Çizelge 3.6. M r püskülünden elde edilen polifenol oksidaz aktivitesinin 4-metilkatekol substrat nda pH ile de imi Çizelge 3.7. M r püskülünden elde edilen polifenol oksidaz aktivitesinin Katekol
substrat nda s cakl k ile de imi
Çizelge 3.8. M r püskülünden elde edilen polifenol oksidaz aktivitesinin 4-metilkatekol substrat nda s cakl k ile de imi
Çizelge 3.9. M r püskülünden elde edilen polifenol oksidaz enziminin termal Denatürasyonu
Çizelge 3.10. M r püskülünden elde edilen polifenol oksidaz aktivitesine karbonhidratlar n etkisi
Çizelge 3.11. M r püskülünden elde edilen polifenol oksidaz aktivitesine inhibitörlerin etkisi 3 5 8 8 28 29 29 30 31 31 32 33 34 35 35
VII
EK L L STES
ekil No Sayfa ekil 1.1. r Püskülü (Stylus maydus)
ekil 1.2. Hücre Parçalanmas ekil 1.3. Amonyum sülfat çöktürme ekil 1.4. Diyaliz
ekil 1.5. Ultrafiltrasyon ekil 1.6. Elektroforez ekil 3.1. r püskülünden elde edilen PFO enziminin
safla rma yöntemleriyle safla rma miktarlar
ekil 3.2. Spektofotometrik ölçümler
ekil 3.3. r püskülünden elde edilen PFO molekül a rl 10 12 13 14 14 16 27 28 39
VIII
Simge Ad Birimi PFO Polifenoloksidaz enzimi -
E.C Enzim kod numaras -
E.Ü Enzim ünitesi E.Ü
Vo Ba lang ç an ndaki h EÜ/dak mL
Vmax Enzimin substrata doydu u andaki h EÜ/dak mL
Km Maksimum h n yar na eri ildi i andaki mol/L
substrat konsantrasyonu
[S] Substrat konsantrasyonu mol/L
[I] nhibitör konsantrasyonu mol/L
V Süpernatant hacmi mL
PEG Polietilen glikol
Nurten ASLAN
1 1.
1.1. Enzimler
Enzimler, canl organizmalardaki biyokimyasal reaksiyonlar katalizleyen ve protein yap nda olan biyolojik aktiviteye sahip moleküllerdir. Canl lar, binlerce çe it enzim ihtiva etmeleri ve hayatsal olaylar bu enzimler sayesinde düzenlemek suretiyle ya amlar devam ettirmektedirler. Enzimler, proteinlerin en büyük ve en özelle mi grubunu te kil eder (Nelson ve Cox 2005).
Biyokimya tarihçesinin ço u enzim ara rmalar n tarihçesidir. Biyolojik kataliz, ilk kez 1700’lerin sonlar nda mide salg vas tas yla etin sindirilmesinin ve 1800’lü y llarda tükrük ve çe itli bitki ekstratlar vas tas yla ni astan n ekere dönü türülmesinin ara lmas çal malar nda tan nm ve tan mlanm r. 1850’lerde Lous Pasteur ekerin maya vas tas yla alkole fermentasyonunun ‘fermentler’ arac yla katalizlendi i sonucuna varm r. 1878’de Frederick W. Kühne bu moleküllere (fermentlere) ‘enzim’ ad vermi ve bu fermentlerin canl maya hücrelerinin yap ndan ayr lmaz oldu unu kabul etmi tir. Bu görü uzun y llar geçerli olmu tur. Daha sonra 1897’de Eduard Buchner maya ekstratlar n ekeri alkole fermente edebildi ini ke fetmi ve fermentasyonu sa layan moleküllerin canl hücre yap ndan
kar ld nda da fonksiyonuna devam etti ini ispat etmi tir. Bu ispat biyokimyac lar , yeni enzimlerin izolasyonu ve onlar n katalitik özelliklerini incelemeleri için te vik etmi tir (Nelson ve Cox.2005).Enzimlerin katalize etti i reaksiyonlar n h zlar , katalize edilmeyen ayn reaksiyonun h ndan 103- 108 kat daha fazlad r. Enzimler o kadar stereospesifiktirler ki, tek bir metabolitin tek bir reaksiyonunu katalize ettikleri gibi, sadece enantiyomerleri de il identik atomlar veya gruplar da parçalayabilirler (Bay u Sözbilir ve Bay u, 2008). Enzimatik reaksiyonlar n h zlar ; s cakl k, pH, zaman, substrat konsantrasiyonu, enzim konsantrasyonu ve inhibitör gibi faktörler etkilemektedir.
1.1.2. Enzim Aktivitesini Etkileyen Faktörler a. Enzim Konsantrasyonunun Etkisi
Enzim reaksiyonunun h , genel olarak, enzim konsantrasyonu ile orant r. Substrat konsantrasyonu sabit tutulup enzim konsantrasyonu artt rsa substrat n tamam enzim-substrat kompleksi olu turana kadar reaksiyon h artar.
b. Substrat Konsantrasyonunun Etkisi
Sabit enzim konsantrasyonunda, enzim reaksiyonunun h belirli bir noktaya kadar substrat konsantrasyonu ile artar. Bundan sonra substrat konsantrasyonunun artmas ile reaksiyon h de mez.
2 c. S cakl n Etkisi
Enzim reaksiyonlar n h s cakl k ile artar. S cakl n her 10°C artmas ile reaksiyon h ortalama iki kat artar. n vitro enzim reaksiyonlar ço u zaman 37 – 40°C’de yap r. Bu s cakl kta reaksiyon h oda s cakl ndakine oranla dört kat daha fazlad r. Fakat belirli bir s cakl k a ld ktan sonra, enzimler denatüre olurlar ve etkilerini kaybederler. Her enzim için birim zamanda substrat en fazla de ikli e u ratt belirli bir s cakl k vard r. Bu
cakl a o enzimin optimum s cakl denir.
d. pH Etkisi
Enzimin reaksiyon h ortam n pH’s na ba r. Belirli bir pH alan nda enzimin etkisi en fazlad r. Bu pH’ya enzimin optimum pH’s denir. Optimum pH’n n a nda ya da yukar nda reaksiyon h daha azd r. Belirli bir pH’dan sonra da enzim tamamen etkisiz kal r.
e. Zaman n etkisi
Yap lan ara rmalar enzimlerin optimum s cakl klar n ve optimum pH’lar n zamana ba oldu unu göstermektedir.
f. Reaksiyon ürünlerinin etkisi
In vitro enzim reaksiyonu devam ettikçe reaksiyon ürünleri enzimi inhibe ettikleri için enzim reaksiyonun h azal r. Bu inhibisyona sebep, reaksiyon ürünlerinin molekül yap bak ndan substrat and rmalar ve enzime substrattan daha fazla ba lanmalar r. Reaksiyon ürünü, enzim proteini ile substrat n birle ti i yerin d nda bir yerde birle ebilirler. Allosterik yere ba lanan madde, enzimin etkili bölgesinde biçimsel bir de iklik gösterebilir. Bunun sonucu olarak, enzim proteininin eklinin bozulmas dolay yla enzimin substrat ile birle mesi substrat n yap na, konsantrasyonuna veya di er faktörlere ba olarak inhibisyona neden olur. Bu etkiye de allosterik etki denir (Seriner 2010).
Enzimler kullan m amaçlar na göre 3 grupta toplanabilir:
1-Üretime yönelik çe itli reaksiyonlar n katalizinde kullan lan safl k dereceleri dü ük olan endüstriyel enzimler,
2-Bilimsel ara rma ve analitik amaçla kullan lan safl k dereceleri yüksek analitik enzimler,
3-Çe itli fizyolojik hastal klar n tedavisinde kullan lan ve safl k derecesi oldukça yüksek klinik enzimler.
Nurten ASLAN
3
1.3. Enzimlerin Kod Numaralar na Göre S fland lmas
1961 y nda enzim komisyonunun haz rlad rapora göre enzimler katalizledikleri reaksiyon tipine göre 6 ana s fa ayr lm ve bu s flarda yer alan her enzim 4 rakamdan olu an bir enzim kod numaras (EC no) ile ifade edilmi tir. EC numaras n ilk rakam enzimin alt ana s ftan hangisinde yer ald , ikinci rakam etki etti i kimyasal yap ve fonksiyonel grubu, üçüncü rakam al grubu (akseptör) ve dördüncü rakam ise o serideki s ra numaras ifade eder (Çizelge 1.1) (Kuzu 2008).
Çizelge 1.1. Enzimlerin kod numaralar na göre s fland lmas
1. Oksidoredüktazlar Redoks reaksiyonlar katalizlerler
1.1. CH-OH grubuna etki edenler 1.2 C-O grubuna etki edenler 1.3 C-CH grubuna etki edenler 1.4 C-NH2 grubuna etki edenler 1.5 CH-NH grubuna etki edenler
2. Transferazlar Fonksiyonel gruplar n transferinde görev yaparlar
2.1 C1- gruplar transfer edenler 2.2 Karbonil gruplar transfer edenler 2.3 Açil gruplar transfer edenler 2.4 Glikozil gruplar transfer edenler 2. 5 N- içeren gruplar transfer edenler 2.6 Fosfat gruplar transfer edenler 2.7 S- içeren gruplar transfer edenler
3.Hidrolazlar Hidroliz reaksiyonlar katalizlerler
3.1 Esterleri hidrolizleyenler
3.2 Glikozid ba lar hidrolizleyenler 3.4 Peptid ba lar hidrolizleyenler 3.5 Di er C-N ba lar hidrolizleyenler 3.6 Asit anhidridlerini hidrolizleyenler
4.Liyazlar Çifte ba a kat lma ve çifte ba n olu um reaksiyonlar
katalizlerler
4.1 C-C liyazlar 4.2 C-O liyazlar 4.3 C-N liyazlar
5. zomerazlar zomerle me reaksiyonlar katalizlerler
5.1 Rasemazlar
4 5.4 ntramoleküler transferazlar
6.Ligazlar Sentez reaksiyonlar katalizlerler
6.1 C-O ba olu turanlar 6.2 C-S ba olu turanlar 6.3 C-N ba olu turanlar 6.4 C-C ba olu turanlar
1.1.4. Oksidoredüktazlar
Oksidoredüktazlar, oksidasyon (yükseltgenme) ve redüksiyon (indirgenme) reaksiyonlar katalizleyen enzimlerdir. Yükseltgenme; moleküle oksijen eklemekle veya molekülden hidrojen ayr lmakla, böylece + de erin artmas yla olur. ndirgenme ise; molekülden oksijen ayr lmakla veya moleküle hidrojen eklemekle yani + de erin azalmas yla olur. Bu tepkimeleri düzenleyen enzimler 2 gruba ayr lm r. 1)Dehidrogenaz: Hidrojen ta yan ve aktaran enzimlerdir. 2)Oksidazlar: Elektron kaybeden ve aktaran enzimlerdir (Nelson ve Cox 2004, Megep 2007)
1.2. Polifenoloksidaz
Polifenoloksidaz (PFO) (E.C.1.14.18.1) yap nda kofaktör olarak bak r içeren oksidoredüktaz s na ait bifonksiyonel bir enzimdir (Turan 2005). Polifenoloksidazlar olarak adland lan enzimler, bakterilerden memelilere kadar tüm filogenetik skalada geni yay m gösteren bak r içeren proteinlerin bir grubudur. Bu grubun ortak özelli i, moleküler oksijen varl nda fenollerin oksidasyonunu katalizleme kapasitelerinin olmas r (Shi ve ark. 2002).
Polifenol oksidazlar (PFO) bak r içeren yayg n enzimlerdir. Bu enzimler fenolik bile ikleri kinonlara oksitler ve bu reaksiyonda moleküler oksijeni kullan rlar. PFO’lar taraf ndan üretilen kinonlar yüksek derecede reaktiftirler ve kendileri biraraya gelip ba lanarak ve ayr ca proteinleri çapraz ba layarak veya alkilleyerek hasar görmü bitki dokular nda ve ekstraktlar nda genellikle gözlenen kahverengi pigmentlerin olu mas na neden olurlar (Constabel ve ark.2008).
Polifenoloksidaz ilk olarak mantarlarda ke fedilen yayg n bir enzimdir. Bunlar bitkide plastidlerin ve kroloplastlar n içinde bulunmas na ra men olgunla bitkinin sitoplazmas nda serbest olarak bulunur, mantarlarda ise genellikle sitoplazmada, baz durumlarda da hücre duvar nda lokalize olmu tur. Sa kl meyve ve sebze dokular nda, PFO enzimlerinin, substratlar olan fenolik bile iklerle temas yok denecek kadar azd r. Bunun ba ca nedeni, enzim ve substratlar n bitkisel hücrenin farkl k mlar nda yer almalar r. Fenolik bile iklerin neredeyse tamam vakuollerde yo unla halde bulunmaktad r. Ancak doku
5
olgunla mas n ileri a amalar nda hücredeki pektinazlar n faaliyetiyle, doku kontrollü ve s rl bir ekilde do al olarak de imlere u rar. Ayr ca, ta ma, hasat ve i leme s ras ndaki etkiler veya uygulanan çe itli i lemlerle hücre ve buna ba olarak doku bütünlü ü bozulmaktad r. Böylece, PFO enzimleri fenolik bile iklerle ve havadaki oksijen ile biraraya gelmektedir (Muchuweti ve ark. 2006).
Bu enzim, bitkinin mikrobiyal ve viral enfeksiyonlara, kötü iklim ko ullar na kar direnci aç ndan önemlidir. Ayr ca PFO'nun faaliyeti sonucu suda çözünmeyen polimerler olu makta bu da zedelenmi dokularda olu abilecek enfeksiyonlar n yay lmas önlemektedir (Turan 2005), (Mayer 2006).
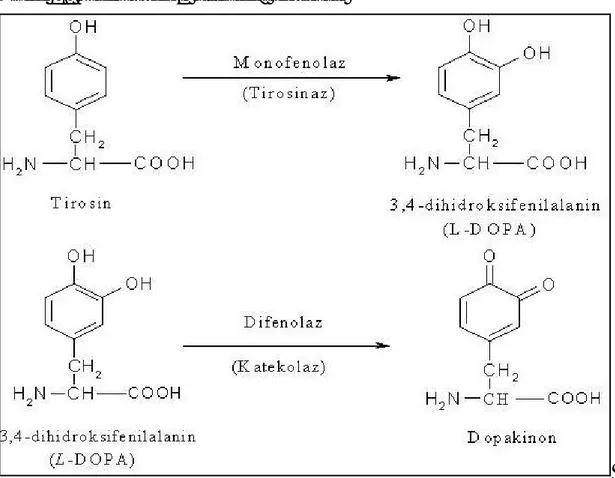
Bak r içeren bir enzim olan PFO iki farkl reaksiyon katalizler ( Zawistowski ve ark. 1991).
1) monofenollerin o-difenollere hidroksilasyonu 2) o-difenollerin o-kinonlara dehidrojenasyonu
Çizelge 1.2. Polifenoloksidaz n katalizledi i reaksiyonlar
Birçok bitkide ve meyvede enzimatik kararma ad yla bilinen renk de imleri gözlenmektedir. Meydana gelen bu renk de imlerinin nedeni enzimatik ve enzimatik olmayan
6
reaksiyonlard r. Enzimatik kararmaya neden olan ba ca enzim polifenoloksidazd r (Do an ve ark. 2003). Meyve ve sebzelerin kararmas , Polifenoloksidaz n katalizledi i fenolik bile iklerin kinonlara, onlar n da daha sonra kondanse renkli pigmentlere oksidasyonu sonucunda ortaya kmaktad r (Lee ve Park 2005). PFO’lar taraf ndan üretilen kinonlar yüksek derecede reaktiftirler ve kendileri biraraya gelip ba lanarak ve ayr ca proteinleri çapraz ba layarak veya alkilleyerek hasar görmü bitki dokular nda ve ekstraktlar nda genellikle gözlenen kahverengi pigmentlerin olu mas na neden olurlar (Constabel ve Barbehenn 2008). Renkli bak r bile iklerinin polimerize olmas yla meydana gelen kinonlar, besin kalitesinin, meyve ve sebzelerin ticari de erlerinin dü mesine sebep olmaktad r (Do an ve ark. 2006). Enzimatik kararman n derecesi fenolik bile iklerin miktar na, da as na, oksijenin varl na, indirgen maddeye, metalik iyonlara, pH'ya, s cakl a ve polifenoloksidaza ba r. Enzimatik kararma
letmeciler için önemli ekonomik bir problemdir (Ertürk 2002).
1.2.1. Polifenol oksidaz n önemi
G da endüstrisinde enzimler son y llarda cazip hale gelmi tir. Yüksek besin de erine sahip g dalar n geli tirilmesi ve üretimi gittikçe önem kazanmaktad r. G dalar n korunmas nda dokulardaki kimyasal olaylar n kontrol alt na al nmas önemlidir. Çünkü özellikle sebze ve meyvelerde meydana gelen kimyasal de imler ürünün renginin bozulmas na ve besin de erinin dü mesine sebep olmaktad r. Bu renk bozulmalar enzimatik ve enzimatik olmayan kimyasal reaksiyonlar sonucu ortaya ç kmaktad r. Çe itli i lemler s ras nda meyve ve sebze dokular herhangi bir ekilde zedelenirse, kesilirse, kabu u soyulursa g dalarda baz istenmeyen kahverengile me reaksiyonlar meydana gelir (Lee ve Park 2005). Sebzelerde ve meyvelerde polifeoloksidazlardan kaynaklanan enzimatik kararma istenmeyen renk, koku ve tad olu umuna neden olmakta, bu da g dalar n besin de erinin önemli ölçüde dü mesine sebep olmaktad r (Do an ve ark. 2003).
1.2.2. Polifenoloksidaz n Tabiattaki Da ve Önemi
PFO ilk kez Schoenbein taraf ndan 1856 y nda yemeklik mantarda bulunmu tur (Öztan 2007). Bitkilerde, mikroorganizmalarda ve özellikle mantar ve baz hayvansal organizmalarda bulunur. PFO'nun varl na kabuklu deniz hayvanlar n baz lar nda da rastlan lm r (Do an 2002). PFO, böceklerde d iskeletin geli mesine yard mc olurken sentezlenen melanin sayesinde mikroorganizmalar hapsederk koruma sa lamaktad r (Sherman ve ark.1991). Memelilerde ise zararl güne nlar na kar koruyucu ajanlar olarak görev yapan melaninlerin biyosentezinden sorumludur (Yoruk ve Marshall 2003). Bitkilerde meydana gelen mekanik zararlara ve virüs, bakteri ve mantarlar taraf ndan olu turulan enfeksiyonlara kar
Nurten ASLAN
7
bitkinin direnç göstermesinde etkili bir role sahiptir (Turan 2005). PFO'nun faaliyeti sonucu olu an kinonlar n ikinci bir polimerizasyon reaksiyonuyla koyu renkli, suda çözünmeyen polimerler olu turdu u bilinmektedir. Polimerlerle doldurulan zedelenmi dokular enfeksiyonun yay lmas na kar bir engel olu turmaktad r (Turan 2005).Baz tah llardan elde edilen undaki yüksek PFO aktivitesinden dolay , ekmek, makarna ürünlerinin kararma gösterdi i enzimatik reaksiyonlar sonucu olu an kinonlar n proteinlerle birle erek onlar n hazmolma yeteneklerini ve bu arada lisin aminoasidinin yararl azaltt bildirilmi tir (Oynan 2000). Çay, kakao ve kahvenin üretiminde enzimatik kararma istenilen önemli biri lemdir. Siyah çay n üretimi süresince meydana gelen biyokimyasal de imler çay n lezzetini veren teaflavin ve tearubinlerin üretilmesi aç ndan önemlidir ( Keskin ve H.Erkmen 1987).
Siyah çay üretiminde enzimatik kararma istenen bir olayd r. Kinon, çaya içim tad ve kalitesini kazand ran teaflavinin ön maddesidir. Bu nedenle çay PFO enzimi üzerinde çok çal ma yap lm r (Gregory and Bendall 1966).
PFO’nun kullan ld bir baska önemli alan ise Parkinson hastal n tedavisinde kullan lan L-DOPA’n n üretimidir (Whitaker ve ark. 1995).
1.2.3. Polifenoloksidaz n Substratlar
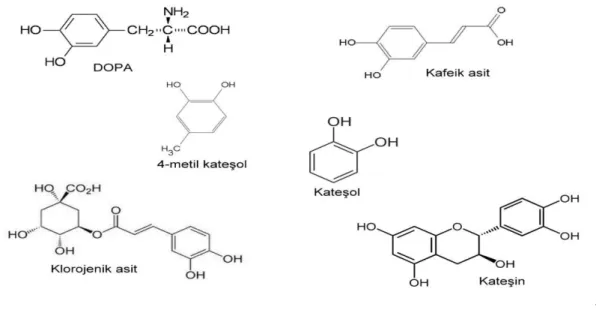
Meyve ve sebzeler çok miktarda fenolik madde içerirler. Bunlardan baz lar polifenoloksidaz n substratlar r. Polifenoloksidaz n sebze ve meyvelerdeki en yayg n do al substratlar , flavonoid tipi fenollerle, basit fenollerdir. Flavonoidler, bitkilerin yenilebilir k sm ile kök, gövde, yaprak, meyve ve tohum k mlar nda bulunur. Bitkilerin çe itli bölümlerinde homojen bir ekilde da lm lard r. De ik konsantrasyonlardaki polifenoller, kararma reaksiyonlar n meydana gelmesinde etkilidir. Bunlardan baz lar katekinler, sinamik asit esterleri, 3,4 - dihidroksifenil alenin (DOPA) 3,4-dihidroksifenil etilamin (dopamin) ve tirozindir (Do an 2002).
Fenolik substratlardan, her bitkide bulunan tirozin ayn zamanda proteinlerin yap olu turan aminoasitlerden biridir. Dopamin (3,4-dihidroksifenil etil amin) ve tirozinin PFO ile hidroksilasyon sonucu olu an L-DOPA (3,4-dihidroksifenil alenin) bitki dokular nda mevcuttur. Katekol, en basit fenoldür. Hidroksilli B halkas na sahiptir (Sakiro lu 1994).
8
Çizelge 1.3. Polifenoloksidaz enziminin baz substratlar n yap
Çizelge 1.4. Fenolik bile ikler
Kaynak Fenolik bile ikler
Elma klorogenik asit (et), katekol, katekin (kabuk), kafeik asit,
3,4-dihidroksifenilalanine (DOPA), 3,4-dihidroksi benzoik asit, p- kresol, 4-methyl katekol, leukosianidin, p- koumarik asit, flavonol glikosides
Kay isoklorogenik asit, kafeik asit, 4-methyl katekol, klorogenic asit, katekin, epikatekin, pirogallol, katekol, flavonols, p- koumarik asit türevleri
Avokado 4-methyl katekol, dopamin, pirogallol, katekol, klorogenic asit, kafeik asit, DOPA
Muz 3,4-dihidroksifeniletilamin (Dopamin), leukodelphinidin, leukosiyanidin
Kakao katekinler, leukoantosiyanidinler, anthosiyaninler, kompleks taninler
Kahve çekirdekleri klorogenic asit, kafeik asit
Patl can klorogenik asit, kafeik asit, komarik asit, sinamik asit türevleri
Üzüm catechin, klorogenik asit, catechol, kafeik asit, DOPA, taninler, flavonoller, protocatechuic asit, resorsinol, hydroquinone, fenol
Salata tirozin, kafeik asit, klorogenik asit türevleri
Istakoz Tirozin
Mango dopamine-HCl, 4-methyl catechol, kafeik asit, catechol, catechin, klorogenik asit, tirozin, DOPA, p- cresol
Mantar tirozin, catechol, DOPA, dopamine, adrenaline, noradrenaline
eftali klorogenik asit, pyrogallol, 4-methyl catechol, catechol, kafeik asit, gallic asit, catechin, dopamine
Armut klorogenik asit, catechol, catechin, kafeik asit, DOPA, 3,4-dihydroxy benzoic asit, p- cresol
Erik klorogenik asit, catechin, kafeik asit, catechol, DOPA
Patates klorogenik asit, kafeik asit, catechol, DOPA, p- cresol, p- hydroxyphenyl propionic asit, p- hydroxyphenyl pirüvik asit, m- cresol
Karides Tirozin
Tatl patates klorogenik asit, kafeik asit, caffeylamide
Nurten ASLAN
9 1.3. Esmerle me Reaksiyonlar :
1.3.1. Kimyasal Esmerle me Reaksiyonlar
Besinlerde enzimatik olmayan yolla renk esmerle mesi; askorbik asit oksidasyonu, karamelizasyon ve Maillard tepkimesi olmak üzere ba ca üç farkl ekildedir (Friedman 1996). 1.Askorbik asit oksidasyonu: Bu asidi içeren meyve ve sebzelerin de ik g dalara lenmesi s ras nda renk esmerle mesine yol açan bir tepkimedir. Bu tepkimede askorbik asit dehidro-askorbik asit ve diketo-gulonik asit üzerinden furfural ve karbon diokside parçalanmaktad r (URL–2 2008).
2.Karamelizasyon: Karbohidratlar n yüksek s cakl klara maruz b rak ld klar nda olu an reaksiyonlar n genel ismidir. Bu reaksiyonlar genellikle ekerlemecilikte ve hamur tatl lar nda kullan lan karamellerin ve geleneksel sukroz uruplar n haz rlanmas esnas nda olu ur.
3.Maillard reaksiyonu: Amino grubu ve indirgen ekerler aras nda genellikle yüksek cakl klarda meydana gelen karma k reaksiyonlar serisidir.
1.3.2. Enzimatik Esmerle me Reaksiyonlar
Enzimatik kararma reaksiyonlar iki ad mda olu ur. Birinci ad m, polifenol oksidaz n(PFO) katalizledi i reaksiyon ile monofenollerin ya da difenollerin o-kinonlara enzimatik oksidasyonu, ikinci ad m ise enzimatik oksidasyon sonucu olu an o-kinonlar n, enzimatik olmayan kondenzasyon ya da polimerizasyon reaksiyonlar sonucu melanin bile iklerini olu turmas r. PFO substratlar (fenolikler ve oksijen) ile temas kurar ( Yörük ve Marshall 2003).
1.4. M r Bitkisi ( Zea mays ) Alem : Plantae ( Bitkiler )
Bölüm :Magnoliophyta ( Kapal Tohumlular ) S f : Liliopsida ( Bir Çenekliler ) Tak m : Poales
Familya : Poaceae(Bu daygiller) Cins : Zea
Tür : Zea Mays Alt türler
Zea mays ''m r'' olarak bilinen Poaceae familyas na ait önemli bir bitkidir. Genellikle çok nemli iklim bölgelerinde yeti tirilebilen tek y ll k özellikle ya doymam ya grubunda olan bir tar m bitkisidir.
10
alt ndad r. Tanesindeki ham ya yulaftan sonra en yüksek de er veren besin maddesidir. Meksika ve Orta Amerika kökenlidir. S fland rma koçan ekli, tane, somak rengine bak larak yap r.(http://tr.wikipedia.org)
1.5. M r Püskülü (Stylus maydis)
r bitkisinin di i çiçekler (koçan püskülü) k sm r. Ortalama 2 m boylar nda, kökü kal n ve sacakl , yapraklar erit gibi uzun, sert ve sivri olan bir bitki olan m n meyvelerinin ucundaki saçaklard r. M r bitkisinde di i organ koçan olarak tabir edilir. Her bir koçanda yakla k 300-1000 adet dane olu ur ve her dane için bir adet püskül bulunur. Koçan püskülü üzerine gelen polen tozlar ile döllenme ba lar. M r püskülü bol miktarda potasyum, sodyum ve kalsiyum içerir. M r püskülünde ayr ca ya , reçine, ekerler ve allontion bulunur. Di i çiçeklerin olgunla mas yla meydana gelen m r tohumlar , tek ve kal n bir sap olan koçan üzerinde düzgün s ralar halinde dizilmi iri taneler eklinde olur. laç olarak kullan lan m r püskülünün içerdi i maddeler ise unlard r: Glikoz ve maltoz gibi ekerler, steroller, reçine, potasyum tuzlar ve uçucu ya .
ekil 1.1. M r Püskülü (Stylus maydis)
Aç k esmer ya da k rm ms renkli hafif ve özel kokusu bulunan m r püskülünün t bbi etkileri ve bunlardan yararlanma yöntemleri öyle özetlenebilir:
1. Sakinle tiricidir.
2. Bedeni güçlendirici toniktir.
1.G ______________________________________________________________________
11 4. drar söktürücüdür.
5. Mesane ta lar dü ürür.
6. Üretrit (idrar yollar enfeksiyonu), sistit (mesane enfeksiyonu) ve prostatit (prostat bezi enfeksiyonu) tedavilerinde etkilidir. Özellikle ayr kotu ve civanperçemi ile birlikte kullan rsa daha etkili olur.
7. Çocuklarda böbrek sorunlar n atlat lmas na yard mc olur.
Bu etkileri sa lamak üzere, m r koçan ndaki di i çiçeklerin döllenme olay gerçekle meden önce ortaya ç kan püskülleri al r. Bunlar kurutuldu unda baz etkilerini yitirdi inden kurutulmadan kullan lmas daha do ru olur. 1 bardak kaynar suyun içine 2 tatl ka kuru ya da taze m r püskülü konur. 10-15 dakika demlendirilerek elde edilen infüzyondan günde iki-üç kez birer bardak içilir (http://www.saglikbilgisi.gen.tr/) (http://tr.wikipedia.org).
1.6. Enzimlerin Safla lma A amalar
Enzimlerin kullan m alanlar her geçen gün artmaktad r. Biyoteknoloji alan ndaki geli meler enzimlerin kullan m alanlar artt rm ve yeni uygulama alanlar n da ortaya kmas ile enzimlerin saf olarak elde edilmesi büyük önem kazanm r. Enzimler bitkisel, hayvansal ve mikrobiyal kaynakl olabilirler. Ticari olarak sat lan enzimler biyolojik kaynaklardan izole edilmektedir(Seriner 2010).
1.6.1. Hücre Parçalanmas
Baz durumlarda bir doku veya hücrenin parçalanarak arzu edilen bir proteinin çözelti içine sokulmas gerekir. Bu amaca ula man n birkaç yolu vard r: tekrarl dondurma ve eritme, sonikasyon, yüksek bas nçla homojenizasyon, filtreleme, veya organik çözücülerle hücre zar n geçirgenle tirilmesi (permeabiliza sonu). Seçilecek yöntem, proteinin ne derece hassas oldu u ve hücrelerin ne derece sa lam oldu una ba r. Bu özütleme sürecinden sonra çözünür proteinler çözelti içine girerler; hücre zar , DNA ve di er hücre bile enlerinden santrifüjleme yoluyla ayr labilirler. Bu süreçte hücrenin proteazlar da çözeltiye kar p ve di er proteinleri sindirmeye ba layabilirler. E er safla lacak protein bu proteazlar taraf ndan parçalanmaya (proteoliz) duyarl ise, bu i lemin h zla tamamlanmas ve özütün so uk tutulmas tercih edilir (http://tr.wikipedia.org).
12
ekil 1.2. Hücre Parçalanmas
1.6.2. Amonyum Sülfat Çöktürmesi
Hücre izolatlar uzakla larak elde edilen ham ekstrakt üzerinden safla rma i lemleri ba lamaktad r. Safla rma i lemlerinin ilk basama çöktürme i lemidir. Proteinler, çözünürlüklerine göre ayr p safla labilirler. Sodyum sülfat veya amonyum sülfat ile çöktürme yöntemleri, proteinlerin ayr lmas nda ve safla lmas nda kullan lan en eski yöntemlerdir. Nötral tuzlar n ilavesiyle yap lan çöktürme proteinlerin fraksiyonlanarak çökeltilmelerinde en çok kullan lan yöntemdir. Ay rma i lemi s ras nda proteinlerin denatüre olmamasi için çöktürme so ukta yap lmal r. Çözünmü proteinlerin amonyum sülfat gibi nötral tuzlar n etkisiyle çökmelerinin nedeni, protein moleküllerindeki ba suyun çekilmesidir. Bu ekilde çöktürülen protein genellikle denatürasyona u ramad için aktivite proteinlerin çözülmesiyle elde edilmektedir. Ayr ca bu tuzlar proteinleri denatürasyon, proteoliz ve bakteriyel kontaminasyona kar korumaktad rlar (Erarslan ve ark. 2000).
1.G ______________________________________________________________________
13
ekil 1.3.Amonyum Sülfat Çöktürme
1.6.3. Diyaliz
Ham ekstrakt n tuz ile muamelesi sonucu olu an çözeltinin santrifüj edilmesiyle elde edilen protein agregatlar çal mada kullan lan uygun tampon ile çözülerek al r.
Elde edilen protein çözeltisinden tuzlar n giderimi veya bir sonraki safla rma i leminin verimli çal mas için çözeltinin tamponunun de tirilmesi s kça ba vurulan durumlardand r. Bu amaçla, genellikle diyaliz i lemi uygulanmaktad r. Protein çözeltisi küçük molekülleri geçiren fakat büyük molekülleri al koyan yar geçirgen bir membrandan olu an diyaliz torbas na konur ve torba çal lan uygun pH’daki tampon içine b rak r. Böylece membrandan tuz ve di er küçük moleküller tampon ile yer de tirirken, protein molekülleri içeride kal r. Moleküllerin bu ekilde geçi i, diyaliz torbas n içi ve tamponun konsantrasyonu e it oluncaya kadar yani ortam dengeleninceye kadar devam eder.
Küçük moleküller ile tuzlar n uzakla lmas ve proteinlerin konsantre edilmesi için uygulanan di er bir yöntem ise, ultrafiltrasyondur (Erarslan ve ark., 2000; Güngör 2008).
14
ekil 1.4. Diyaliz
1.6.4. Ultrafiltrasyon
Diyaliz temelinde çal r; aradaki fark küçük moleküllerin ortamdan ayr lmas için hidrostatik bas nç veya santrifüj gibi kuvvetlerin etkisiyle proteinleri molekül a rl klar na göre diyalize oranla daha h zl ve daha yüksek verimle ayr lmalar sa lamas r.
Nurten ASLAN
15 1.6.5. Jel Filtrasyon Kromatografisi
Molekül büyüklüklerine göre proteinleri ay rmada kullan lan en uygun ve en etkili yöntem, jel filtrasyon kromatografisidir. Jel filtrasyon kromatografisi; farkl molekül büyüklüklerindeki proteinlerin ayr esas alan kromatografidir. Jel filtrasyon kromatografisinin temeli, moleküllerin çözücü ve belirli bir gözenek çap na sahip olan durgun faz aras nda ayr lmas na dayanmaktad r.
Ay rma i lemi kürecik eklinde olan gözenekli matriksin çözücü ile çevrelenmi olarak bir kolona doldurulmas ve üzerinden örneklerin geçirilmesi ile sa lanmaktad r. Kolon matriks malzemesi sefadeks‘, sefaroz‘ gibi bir polisakkarit veya Biogel‘ gibi bir poliakrilamid türevi olabilir. Durgun fazdaki matriksin gözeneklerinden daha küçük ve daha büyük moleküller içeren bir numune dü ünüldü ünde; gözenek büyüklü ünden daha küçük olan moleküller matriksin gözenekleri içine girerler ve kolon boyunca daha yava hareket ederler. Gözenek büyüklü ünden daha büyük olan moleküller ise durgun faz taraf ndan d ar da b rak rlar ve böylelikle kolonu daha önce terk ederler. Ara büyüklükteki moleküller matriks gözenekleri içine girebilirler fakat kolon içinde küçük moleküllere göre daha k sa süre kal rlar. Sonuç olarak, moleküllerin hepsi kolondan azalan büyüklük s ras na göre ayr lmaktad r (Erarslan ve ark. 2000).
1.6.6. Elektroforez
Elektroforez, yüklü moleküllerin elektrik alanda göçüdür. Sulu bir çözelti içinde, süspansiye ya da çözünmü küçük elektrik yüklü parçac klar n, uygulanan bir elektrik alan n etkisi ile göç etmesi sürecine elektroforez denmektedir. Bu küçük parçac klar; hücre organelleri, bakteri hücreleri, virüsler, protein molekülleri veya sentetik parçac klar olabilir. Bir molekülün jeldeki elektroforetik hareketi, yine o molekülün büyüklü üne, yap na, yo unlu una, jelin gözenek aral na, tamponun pH‘s na, iyonik gücüne ve ortam n s cakl na ba r. Farkl göçme h zlar na göre ta materyal üzerinde ayr lan proteinler, boyanarak görünür hale getirilir ve elde edilen protein bantlar , kantitatif olarak de erlendirilir. Elektroforezde kullanilan ta iy materyal; ka t, selüloz asetat tabakas , ni asta jeli, poliakrilamid jeli, agar jeli gibi maddeler olabilir. Ta materyalin çe idine göre de farkl elektroforez yöntemleri tan mlan r (Gül Güven 2007).
16
ekil 1.6. Elektroforez
1.7. nhibisyon
Enzim inhibisyonu, enzimin katalitik ya da düzenleyici merkezleri olarak tan mlanan aktif bölgelerine spesifik olarak ba lanan inhibitörler ile enzim aktivitesinin azalt lmas olarak tan mlanabilir. nhibitörler genellikle küçük molekül a rl na sahip bile ikler ve iyonlard r.
Bir inhibitörün enzime ba lanma ekli elde edilmi deneysel verilerin de erlendirilmesi için büyük bir öneme sahiptir. nhibitörlerün ço u enzime tersinir olarak ba land klar için bunlar enzimden tekrar ayr labilirler. Bununla birlikte baz inhibitörler enzime çok kuvvetli bir ekilde (kovalent ba larla) ba lanabilirler ve bu inhibitörler enzimden ayr lmazlar.Bir çok inhibitör yap sal olarak enzimin substrat na benzeyen maddelerdirler. Bu maddeler substratla kar la ld nda ya hiç reaksiyon vermezler ya da çok yava reaksiyon verirler. Enzim inhibitörlerinin etkin olarak rol oynad çe itli mekanizmalar vard r. Enzim, substrat ve inhibitör aras nda çe itli inhibisyon mekanizmalar vard r.
1.8. Polifenoloksidaz le lgili Çal malar n Özeti
Literatür taramas s ras nda polifenoloksidazla yap lm çok say da çal maya rastlan lm r.
Nurten ASLAN
17
Aydemir (2004) taraf ndan yap lan çal mada, enginardan (Cynara scolymus L.) elde edilen ekstraktta oldukça yüksek PFO aktivitesi bulundu u rapor edilmi tir. Enginardan elde edilen enzimin pH 5–7 aral nda en yüksek aktivite gösterdi i ve pH 6–7 aral nda 25 C’de 60 dak. inkübasyon sonunda aktivitesini korudu u belirtilmi tir. Enzim için en etkin inhibitörün ise dithiotheritol oldu u tespit edilmi tir.
Gülçin ve ark. (2005), rgan otunda PFO enzimini amonyum sülfat çöktürmesi, diyaliz ve CM Sephadex iyon de im kromato rafisi arac yla safla rm ve karakterize etmi tir. PFO’nun katekol, 4-metil katekol, pirogallol, L-tirozin, kate in ve transsinnamik asite kar aktivitesini belirlemi ler, en iyi substrat n L-tirozin oldu unu bildirmi lerdir. Ayr ca çe itli inhibitörlerin etkileri test edilmi ve ditiyokarbomat en etkili inhibitör olarak bulunmu tur.
Gawlik-Dziki ve ark. (2007), brokoliden elde edilen PFO'nun karakterizasyonunu incelemi ler ve en iyi aktivitenin katekol ve 4-metilkatekol substratlar için pH 5.7'de oldu unu ve en etkili inhibiörün ise sodyum sülfat oldu unu belirlemi ler.
akiro lu ve ark. (2008), dereotundan polifenol oksidaz enzimini izole ederek çe itli kinetik özelliklerini incelemi tir. Katekol, klorogenik asit ve dopamin substratlar için Km ve Vmax de erleri belirlenmi tir. nhibitör çal malar nda substrat olarak katekol kullan larak L-sistein klorür, askorbik asit ve glutatyon ile çal lm r. En etkili inhibitörün L-sistein klorür oldu u belirlenmi tir.
Cai ve ark.( 2015) lotus tohumlar ndan polifenol oksidaz enzimini amonyum sülfat ve afinite kromatografisi ile safla rm lar. Molekül a rl 38,6 kDa. , optimum s cakl 20o C , optimum Ph’ y 7 olaak tespit etmi ler. Katekol subsrat için Km 6.04 mM Vmax’ ise 416,67 U olarak bulmu lar.
Cheema ve Sommerhalter (2015) mangodan ham eksrakta PFO nun özelliklerini ara rm lar. PFO aktivitesini pyrogallol, 3-metilkatekol, katekol, gallic asit, ve protocatechuic asit kullanarak optimum pH aral pH 5.4 ve 6.4, km de erlerini ise 0.84 -4.6 mM aral nda tespit etmi ler. Enzimin molekül a rl ise 53, 112, and 144 kDa oldu unu belirtmi ler.
Mdluli (2005), Afrika’da yeti en Scalerocarya birrea subsp. caffra’den elde ettigi PFO ve peroksidaz safla rarak karakterizasyonunu belirlemeye çal r.
Mozetic ve ark. (2006), Rebula üzümlerinden elde ettikleri polifenoleri Folin–Ciocalteu metoduna göre analiz etmi lerdir.
Mayende ve ark. (2006), yapraklardan izole ettikleri termofilik Basillus spp. taksonlar ndan elde ettikleri PFO’yu incelemi lerdir.
Thipyapong ve ark. (2004), fotosentez s ras nda ortaya stres durumunda yükselen PFO aktivitesini domateste inhibe etmeye çal lard r.
18
Onsa ve ark. (2000), Hint irmi inden PFO'yu k smen safla rarak baz kinetik özelliklerini incelemi ler.
Ayaz ve ark. (2008), Döngel (Mespilus germanica L.) meyvelerinden elde edilen PFO enzimi için 4-metilkatekol’ün en iyi substrat oldu unu, optimum pH’n n 7 ve optimum
cakl n 30 C oldu unu belirlemi ler. Km de erini ise 26 mM olarak tespit etmi ler. Sodyum metabisülfitin en iyi inhibitör oldu unu da rapor etmi ler.
Arslan ve ark.(1998), Malatya kay ndan (NH4)SO4 çöktürmesi ve diyalizle k smi safla lan PFO enziminin katekol, L-dopa ve gallik asit gibi difenoliklere kar aktivite gösterdi i belirlenmi tir. Enzimin en iyi aktiviteyi katekol substrat na kar gösterdi i, bu substrat için Km de erinin 6,6 mM ve enzimin optimum pH’s n ise 8,5 oldu u belirlenmi tir. Askorbik asit, 2-Merkapto etanol, sodyum meta bisülfit ve tiyoürenin enzim aktivitesini inhibe etti i gösterilmi tir.
Ünal (2007), Anamur muzundan polifenol oksidaz enzimini safla p, enzimin karakteristik özelliklerini belirlemi tir. PFO‘nun muz için optimum s cakl 30°C bulunmu tur. Enzimin optimum pH derecesi 7.0 olarak bulunmu tur. nhibitör testlerinde ise askorbik asit ve sodyum metabisülfit en etkili inhibitör olarak tespit edilmi tir.
Yemenicio lu ve ark. (1998) , yapt klar çal mada Hale Haven eftalilerinde polifenol oksidaz enziminin niteliklerini ara rm lard r. Ham, yar olgun ve tam olgun eftalilerden öncelikle aseton tozu haz rlanm r. Aseton tozundan haz rlanan enzim çözeltisinde optimum pH, Michaelis sabiti, maksimum h z ve enzim aktivitesinin termal inaktivasyonu incelenmi tir. Olgunluk durumuna göre enzimin optimum pH derecesi 6.0-6.5 aras nda bulunmu tur. Tam olgun eftalilerde elde edilmi enzimin pH 6.2 de oldu u saptanm r. Ayr ca Km de erinin 14.3 mM ve Vm de erinin 1.25 Abs.dk/mL oldu u tespit edilmi tir. Enzimin termal inaktivasyon kineti i 70°C‘de ara lm r.
Corsini ve ark. (1992), patatesten (Solanum tuberosum) safla rarak elde ettikleri PFO enziminin aktivitesi üzerine yapt klar çal malarda enzim aktivitesinin patatesin yumru, kök ve çiçeklerinde oldukça yüksek olmas na ra men yaprak ve saplar nda daha dü ük seviyelerde oldu unu tespit etmi lerdir. PFO enzimi tarafindan meydana gelen enzimatik esmerle menin özellikle patatesin yumrular nda daha fazla oldu unu rapor etmi lerdir.
Kim ve Kim (2013) , Jinseng köklerinden polifenol oksidaz enzimini karboksimetil sefaroz ve fenil sefaroz kromatografisini kullanarak safla rm lar. Moleküler a rl n SDS PAGE ile 40kDa. oldu unu ve en iyi subsrat n ise Pyrogallol oldu unu tespit etmi ler.
Altunkaya ve Gökmen (2013), durum bu day ndan iyon de im kromatografisi ile safla rd klar polifenol oksidaz enziminin optimum s cakl 40 C ve optimum Ph s 6.5 oldu unu ve 26.33 kat safla rd klar rapor etmi lerdir.
Nurten ASLAN
19
Orozco ve ark.(2011), Pouteria sapota bitkisinden iyon de im kromatografisi ile safla rd klar polifenol oksidaz enziminin optimum s cakl 35o C ve optimum Ph’s n 7 oldu unu ve en uygun subsrat n piyrogallol oldu unu ve askorbik asit ve sodyum metabisülfitin etkili inhibitör oldu unu rapor etmi lerdir.
Nurten ASLAN
21 2. MATERYAL VE METOT
2.1. Materyal
2.1.1. Çal mada Kullan lan Bitki Örneklerinin Temini ve Analize Haz rlanmas Bu çal mada kullan lan Zea mays Diyarbak r'da yerel bir marketten temin edildi. Al nan m rlar n kabu u soyulmadan çok k sa sürede laboratuvara getirildi. Kabuklar soyulduktan sonra püsküller topland ve saf suyla y kand .
2.1.2. Çal mada Kullan lan Kimyasal Maddeler
Enzimin safla lmas ve biyokimyasal özelliklerinin belirlenmesinde kullan lan katekol, askorbik asit Merck (Darmstadt, Almanya) firmas ndan temin edilmi tir. PEG (polietilen glikol), amonyum sülfat, , NaH2PO4, 4-metilkatekol, sukroz, fruktoz, riboz, glukoz akrilamid / bis akrilamid, Sodyum azid,SDS, EDTA ,Sephadex G-100 jel filtratsyon resin ve serum albumin Sigma-Aldrich (St. Louis, Amerika) firmas ndan temin edilmi tir.
2.1.3. Çal mada Kullan lan Cihazlar -Su Banyosu : Memmert
-Spektofotometre :Thermo scientific ( Genesys 105 UV V15) - Varion -Hassas Terazi : Denver ( Instrument )- Gec-Avery
-Magnetik Kar : Dragon lab. MS - Heidolph MR Hei- Standart -pH metre : Hanna
-Ev tipi Blender : Arzum -Santrifüj Nüve : NF-200
-2 adet Buzdolab : Vestel dondurucu- Arçelik dondurucu -Vortex :Grant bio (pv-1)
-Micropipet : soterm- B OHit - Eppendorf
-Elektroforez : Mini Protean cell (Bio-RAD) -Wealtec biolab elite 300 Etüv :Memmert
2.2. Ham Enzim Özütünün Haz rlanmas
Al nan m n kabuklar soyulup püskülleri topland ktan sonra hassas terazide 14.45 gr olarak tart ld . M rlar n püskülleri topland ktan sonra saf suyla y kand . 10 mM askorbik asit, %4 PEG içeren 100 ml 0.1 M, pH 6.5 fosfat tamponunda homojenize edildi. Elde edilen homojenat, s k gözenekleri olan ve birkaç kez katlanarak haz rlanan beyaz bir tülbent yard yla filtre edildi. Homojenat daha sonra 10.000 x g’de 30 dakika +4ºC’de santrifüjlendi. Santrifüj sonras , berrak k sma % 80 amonyum sülfat çökeltmesi uyguland . Çökeltilen
22
fraksiyonlar 10.000 x g’de 30 dakika, + 4ºC’de santrifüj edilerek ayr lm r. Çökelti, az miktarda 0.01 M pH 6.5 fosfat tamponunda çözülmü ve +4ºC’de ayn tampona kar bir gece boyunca diyaliz edildi.
2.3. Polifenoloksidaz n Safla lmas
Diyaliz sonras elde edilen ekstart %80 amonyum sülfat doygunlu a getirildi. Bunun için haz rlad z kat amonyum sülfat, magnetik kar da, buz banyosu üzerinde olan ham ekstrata yava yava eklendi. Her eklenmede öncekinin tamamen çözünmü olmas na dikkat edildi. Çözünme i lemi bittikten sonra süspansiyon santrifüj tüplerine e it olarak doldurularak 10000 rpm'de 20 dk santrifüj edildi. Üstte kalan s k m at larak geri kalan k m tek bir tüpte toplan ld . 10.5 ml numune ç kt . Numune diyaliz tüpüne dolduruldu. Bir behere b rak lan pH’s 6.5 olan fosfat tamponuyla 24 saat diyalize b rak ld . Dü ük devirde çal an bir magnetik kar üzerinde buzdolab nda gerçekle tirilen i lem sonucu safla lan enzime ultrafiltrasyon uyguland .
2.4. Ultrafiltrasyon
Ultrafiltrasyon için Amicon Ultra 15 (Millipore) ultrafiltrasyon santrifüj tüpleri kullanilmistir. Hücrelerden ayrilmis PFO üretim ortami, ultrafiltrasyon tüplerinde santrifüjlenerek 5 kDa molekül agirligin altindaki moleküllerin ortamdan uzaklastirilmasi saglanmistir. Santrifüj kosullari +4 ºC, 4000 g’de (6200 rpm) 40 dakikadir. Santrifüj sonrasinda filtrenin üzerinde kalan ve yüksek molekül a rl klara sahip proteinleri içeren k mlar toplanm r. Toplanan ultrafiltrasyon ürünü kromatografi a amas na geçilmeden önce, olas partiküllerin uzakla lmas amaciyla +4 ºC, 14000 rpm, 10 dakika ko ullar nda santrifüjlenmi tir (Gül Güven 2007).
2.5. Jel Filtrasyon Kromotografisi
% 80’lik amonyum sülfat çöktürmesi sonucu PFO aktivitesi görülen süpernatant
diyaliz edildikten sonra1,5 mL/min ak oran yla Sephadex G-75 kolonundan (1.2 x 70 cm)geçirildi. Elüsyon i lemi 0,1 M fosfat tamponu (pH=6,5) ile yap ld . 3’er ml’lik
tüpler halinde toplanan elüatlarda 280 nm’de kalitatif protein analizleri yap ld . Protein olan tüplerde enzim aktiviteleri 420 nm’de belirlendi.
2.6. Nativ PAGE ile Elektroforetik Analiz
Safla rman n çe itli a amalar ndan elde edilen aktif enzimatik fraksiyonlar ve ham preparasyon (crude extract) Laemmli'nin nativ PAGE elektroforez tekni i ile analiz edildi
Nurten ASLAN
23
(Laemmli, 1970). Nativ PAGE tekni i öyle uyguland ; 4 ºC' de % 7 'lik iki paralel jel kullan larak k smi denatüre artlar alt nda elektroforetik ko madan sonra jellerden biri protein bantlar tespit etmek için Coomassie Brillant Blue R-250 (BioRad) ile boyanarak i lemden geçirildi, di er jel ise baz modifikasyonlarla daha önce tan mlanan metotlar (Mishra et al. 2012) kullan larak PFO aktivitesi için boyand . Aktivite jeli daha sonra 30 mM katekol içeren 0.1 M fosfat tamponu (pH 7.0) de 50 ºC de 100 dak. inkübe edildi. Enzimin varl kahverengi band n olu mas yla tespit edildi. Daha sonra jel % 30’luk metanol içinde bekletildi.
2.7. Enzimin Moleküler A rl n Hesaplanmas
Enzimin molekül a rl , sigmadan al nan (Sigma SDS7B2): alpha 2 -makroglobulin (180 kDa), -galaktosidaz (116 kDa), laktoferrin (90 kDa), piruvat kinaz (58 kDa), fumaraz (48.5 kDa), laktik dehidrogenaz (36.5 kDa), triozfosfat izomeraz (26.6 kDa) gibi moleküler rl k standartlar kullan larak belirlendi (Laemmli 1970). Enzimin ald mesafe semilogaritmik bir skalada standart proteinlerle kar la larak belirlendi.
2.8. Spektrofotometrik Ölçümler
Kinetik i lemler Thermo scientific Genesys 105 UV spektometresi kullan larak gerçekle tirildi. Reaksiyon 1cm'lik k yolu olan 3 ml hacimli kuartz küvet içerisinde gerçekle tirildi. Her ölçümde küvetteki reaksiyon solüsyonun hacmi 3 ml olarak sabit tutuldu. Enzim aktivitesi 420 nm’de 60 sn boyunca absorbanstaki art tan belirlenmi tir. Ölçümler iki paralelli olarak yap lm ve sonuçlar ortalama de erler eklinde ifade edilmi tir. Optimum pH’daki 2.8 ml tampon 0.1 ml substrat çözeltisi 0.1 ml enzim çözeltisi ile kar ld ktan sonra absorbanstaki art otomatik olarak kaydedilmi tir. 1 ünite PFO aktivitesi 25°C’de dakikada 0.001 birimlik absorbans art na neden olan enzim miktar olarak tan mlanm r (Ünal ve
ener 2006).
2.9. Enzim Aktivitesine pH' n Etkisi
Enzimin en yüksek aktivite gösterdi i optimum pH’y bulabilmek için pH 3-10 aral klar nda aktiviteler ölçülmü tür. pH 3-5 için 0.1 M sitrat tamponu, pH 6-10 için 0.1 M fosfat tamponu kullan lm r. PFO aktivitesi farkl tamponlarda, standart reaksiyon kar kullan larak ölçülmü tür. Enzimin en yüksek aktivite gösterdi i pH de eri optimum pH olarak belirlenmi ve di er deneyler bu pH’da gerçekle tirilmi tir.
24 2.10. Enzim Aktivitesine S cakl n Etkisi
r püskülünden elde elilen polifenoloksidaz için substrat olarak katekol ve 4-metilkatekol kullan larak optimum s cakl k çal malar gerçekle tirildi. 20-90 °C aras ndaki farkl s cakl k de erlerinde enzim aktivitesi spektrofotometrik ölçümlerle yap ld . Ölçümlerde kullan lan substrat ve tampon çözeltilerinin s cakl su banyosu yard yla ayarland . Enzim en son eklenerek hemen ölçüme al nd . Reaksiyon kar 2.8 ml tampon, 0.1 ml substrat, 0.05 ml enzim kar ndan meydana geldi.
2.11. Termal naktivasyon
M r püskülünden elde edilen polifenoloksidaz enziminin inaktivasyonunun belirlenmesi için 60°C, 70°C ve 75°C s cakl k de erleri seçildi. Bu amaçla katekol substrat kullan ld . Bu çal ma için deney tüplerine konulan enzim çözeltisi sabit zaman aral klar nda çe itli s cakl klarda inkübe edildi. S cakl n enzim stabilitesine etkisini belirlemek için enzim 60ºC’de (5, 10, ve15 dk), 70ºC’de (5, 10, ve 15 dk) ve 75ºC’de ( 5, 10,15 dk ) ld ktan sonra enzim aktiviteleri belirlenmi tir. nkübasyon süresi sonunda enzim su banyosundan ç kar p buz banyosunda so utulmu , ard ndan oda s cakl na getirildikten sonra enzim aktivitesi standart reaksiyon kar kullan larak ölçülmü tür. Kontrol olarak s cakl a maruz b rak lmayan enzim kullan lm r.
2.12. nhibitörlerin Etkileri
nhibitörlerin enzim aktivitesine etkisini belirleyebilmek için 0.5 ve 1 mM konsantrasyonlarda 3 farkl inhibitör (EDTA,SDS,sodyum azid) kullan larak enzim aktivitesi ölçülmü tür. nhibisyon yüzdesi ise a daki formülden hesaplanm r:
nhibisyon (%) = [(Ao – Ai)/Ao)]*100
Ao: nhibitör kullanmadan belirlenen enzim aktivitesi Ai: nhibitör varl nda enzim aktivitesi.
2.13. Karbonhidratlar n etkisi
Karbonhidratlar n enzim aktivitesine etkisini belirleyebilmek için 0.5 ve 2 mM konsantrasyonlarda 4 farkl karbonhidrat (sukroz, fruktoz, riboz ve glikoz) kullan larak enzim aktivitesi ölçülmü tü.
2.14. Protein çeri i
2.14.1. Kalitatif Protein Tayini
2.MATERYAL VE METOT____________________________________________________
25
fenilalanin ve triptofan) 280 nm’deki UV nlar absorblamalar esas na dayanmaktad r (Segel 1968). Çal mam zda kromatografi a amas nda elde edilen numune örneklerinde protein tayini yap ld .
2.14.2. Lowry Metodu
Enzimlerin protein içeri i standart olarak s r serum albumini kullan lan Lowry metoduna göre belirlenmi tir (Lowry 1951).
Konsantrasyonu bilinen standart protein çözeltisi (1 mg/mL BSA) haz rland . Konsantrasyonunu bilmedi imiz çözeltideki protein miktar Lowry yöntemine göre hesapland . Artan konsantrasyonlarda haz rlanan standart ve 50 L enzim çözeltisi al narak tüplerin hepsine 5 mL alkalin çözeltisi eklendi ve 15 dk 40°C’de bekletildikten sonra tüplerin hepsine 1:1 oran nda seyreltilmi 500 L Folin reaktifi (FCR) eklendi ve 30 dk karanl kta bekletildikten sonra 660 nm’de spektrofotometrede absorbans de erleri ölçüldü.
2.15. Polifenol Oksidaz Enzimine Substrat Konsantrasyonunun Etkisi
Enzimin maksimum h n (Vmax) ve Michaelis-Menten sabitinin (Km) bulunmas için gerçekle tirilen kinetik çal mada çe itli konsantrasyonlarda katekol (1-22 mM), 4-metil katekol (2-18 mM) çözeltileri kullanarak enzim aktivitesi ölçümleri yap lm r. Enzimin Km ve Vmaks de erleri Lineweaver-Burk metoduyla grafiksel olarak hesaplanm r. Enzimin maksimum h ve Km de eri tayin edilirken spektrofotometrik olarak 420 nm’de 60 s aktivitesi izlenmi tir. Daha sonra absorbans-zaman grafi inden ilk h zlar hesaplanm r. Bu ilk h z de erleri MichaelisMenten ([S]’a kar V) ve Lineweaver-Burk grafi inde (1/[S]’ye kar 1/V) yerine konularak Km ve Vmax de erleri bulunmu tur.
Nurten ASLAN
27
3.ARA TIRMA BULGULARI VE TARTI MA 3.1.Enzimin Safla lmas
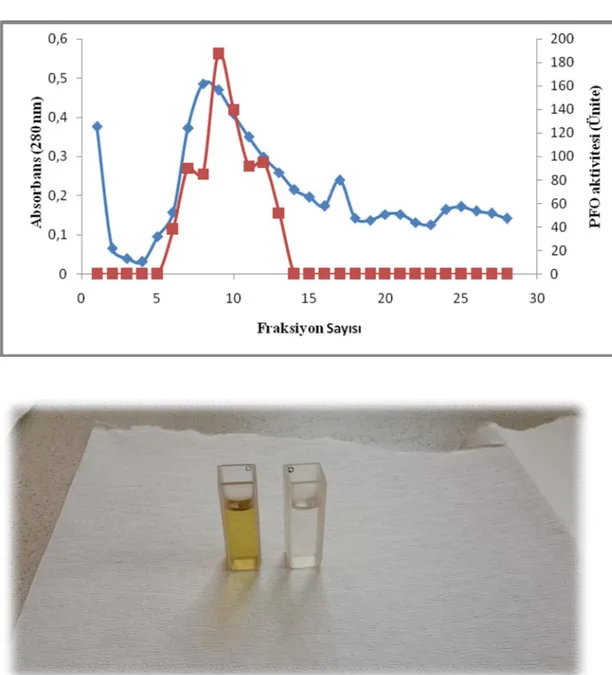
Jel kromatografisinde elde edilen fraksiyonlar n her birinde absorbans (280 nm) ve enzim aktivitesi ölçümleri yap lm r. Aktivite ölçümleri pH’s 4.98 olan 0.1 M fosfat tamponunda haz rlanm 0.1 M katekol kullan larak yap lm r. Toplanan fraksiyonlara kar absorbans ve PFO aktiviteleri Çizelge 3.1’de verilmi tir. Çizelgeden de görüldü ü gibi tek bir aktivite piki elde edilmi . A pikinde 6. Ve 13. fraksiyonlar birle tirilmi ve pikin biyokimyasal özellikleri belirlenmi tir. Safla rma basamaklar ekil 3.2 verilmi tir. ekil 3.2’de görüldü ü gibi enzim 10 kat safla lm r.
ekil 3.1. r püskülünden elde edilen polifenoloksidaz enziminin safla rma yöntemleriyle safla rma miktarlar Safla rma basamaklar Hacim ( ml) Total Protein (mg) Total Ac-Aktivite (U) Spesifik aktivite (U/mg protein) Safla rma (kat) Verim (%) Ham ekstrakt 16 12.43 3280 263.87 1 100 (NH4)2SO4 çöktürme, diyaliz ve ultrafiltrasyon 1.5 3 2311.5 770.5 3 70.4 Sephadex G-75 0.75 0.232 606 2612.06 10 18.47
28
Çizelge 3.1. Polifenoloksidaz enziminin Sephadex G-75 jel geçirgenlik kromatografisi ile elde edilen
fraksiyonlar
ekil 3.2. Spektofotometrik Ölçümler
3.2. Substrat Spesifikli i
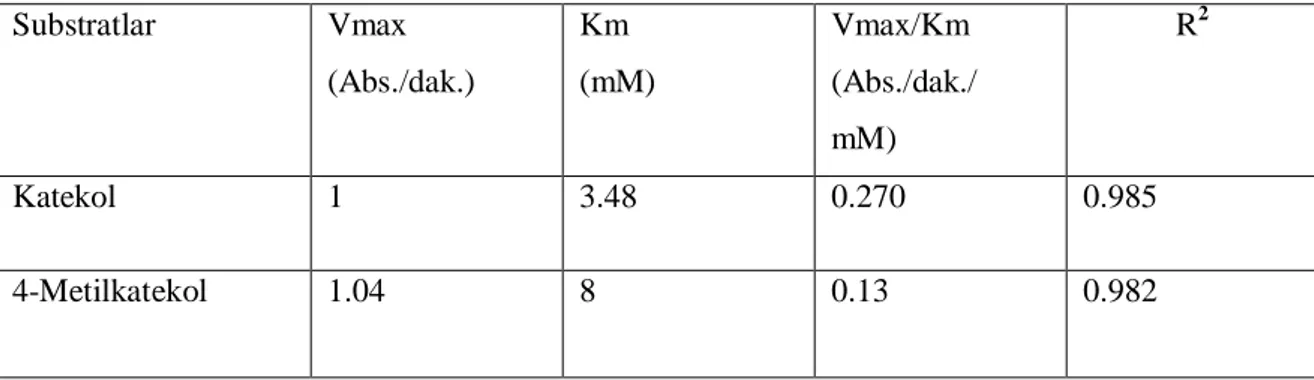
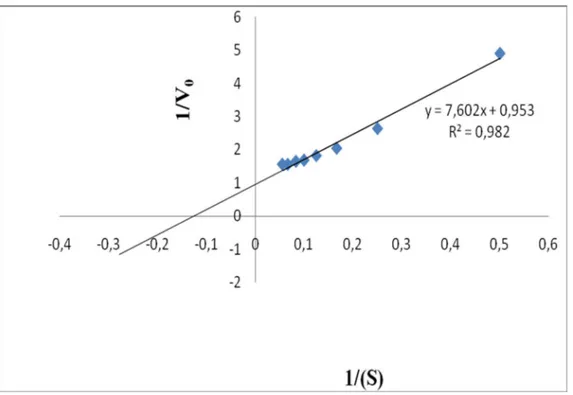
Michealis sabiti (Km) ve maksimum h z (Vmaks)’ belirlemek için substrat olarak çe itli konsantrasyonlarda katekol (1-22 mM), 4-metil katekol (2-18 mM) çözeltileri kullanarak enzim aktivitesi ölçümleri yap lm r. Enzimin Km ve Vmaks de erleri Lineweaver-Burk metoduyla grafiksel olarak hesaplanm r. Elde edilen sonuçlar Çizelge 3.2 gösterilmi tir.
Nurten ASLAN
29
Bir enzim için Km de eri enzimin substrata olan ilgisini gösterir ve bu de er ne kadar küçükse enzimin substrata ilgisi o kadar yüksektir. Km de erleri k yasland nda enzim ilgisinin en yüksek oldu u substrat katekoldür (Km: 3.48 mM). Vmaks/Km oran da enzim için en iyi substrat gösterir. Bu de erin en yüksek oldu u substrat katekoldür (Vmaks/Km : 0.270 ) Literatür taramas yap ld nda Katekol substrat kullan ld nda mamey bitkisinin Km de eri : 44 mM (Palma- Orozco ve ark.2014),kay meyvesinin Km de eri : 6.6 mM olarak tespit edilmi tir. (Arslan ve ark.1998).4-metil katekol subsrat olrak kulan ld nda ise mangonun Km de eri 3.14 mM olarak bulunmu tur (Palma- Orozco ve ark.2014),(Arslan ve ark.1998).
Çizelge 3.2. Polifenoloksidaz aktivitesi için çe itli substratlarla elde edilmi veriler
Substratlar Vmax (Abs./dak.) Km (mM) Vmax/Km (Abs./dak./ mM) R2 Katekol 1 3.48 0.270 0.985 4-Metilkatekol 1.04 8 0.13 0.982
30
Çizelge 3.4. 4-Metil katekol subsrat na ait Lineweaver-Burk grafi i
3.3. Optimum pH
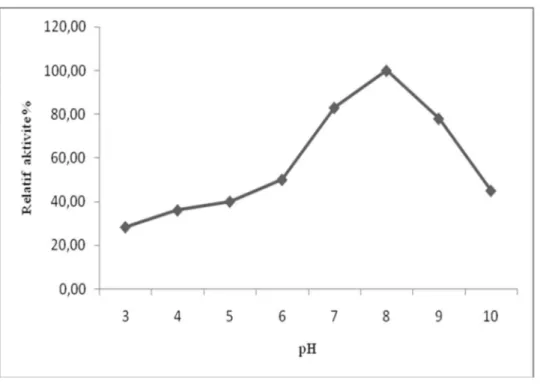
r püskülünden elde edilen polifenoloksidaz n optimum pH de erleri katekol ve 4-metilkatekol substratlar kullan larak belirlenmistir. Elde etti imiz veriler Çizelge 3.5 ve Çizelge 3.6’da gösterilmistir. Optimum pH enzimlerin maksimum aktivite gösterdikleri bir pH veya pH aral vard r. Bu optimum pH’n n alt nda ve üzerinde aktiviteleri dü er. Enzimler kuvvetli asit ve bazlara fazla dayan kl de ildirler. Optimum pH’n n alt nda ve üstündeki pH’larda enzim veya substratta mevcut fonksiyonel gruplar n yap lar nda de meler olu ur ve reaksiyon h da de ime u rar (Koolman ve Roehm, 2005). Katekol kullan larak yap lan çal malarda Amasya elmas n, Yali armutunun ve kakaonun optimum pH s 7.0 olarak tespit edilmi tir. (Oktay ve ark. 1995), (Zhou ve ark.1991), (Lee ve ark. 1991). 4-metil katekol kullan ld nda patl can için pH 6.0 olarak bulunmu tur (Do an ve ark. 2002), (Zhou ve Feng 1991).
3.ARA TIRMA BULGULARI VE TARTI MA______________________________
31
Çizelge 3.5. r püskülünden elde edilen polifenoloksidaz aktivitesinin katekol substrat nda pH ile de imi
Çizelge 3.6. r püskülünden elde edilen polifenoloksidaz aktivitesinin 4-metilkatekol substrat nda pH ile de imi
32 3.4. Optimum S cakl k
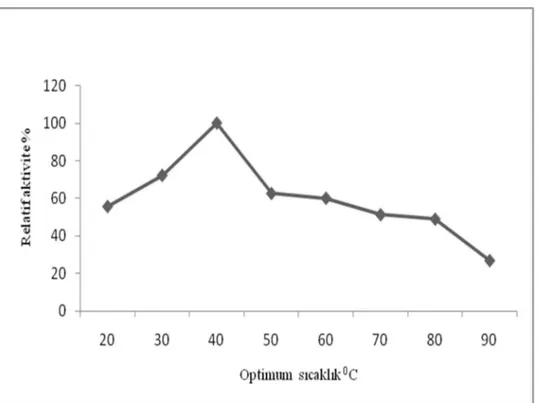
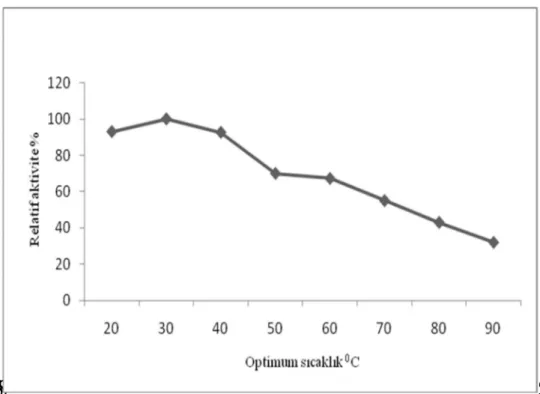
Enzimlerle katalizlenen reaksiyonlar n h zlar da s cakl kla artmakla birlikte, yüksek cakl klarda enzimler protein yap lar ndan dolay aktivitelerini kaybederler. Yüksek s cakl k enzimleri inaktive edebilir. Katekol ve 4-metilkatekol substratlar kullan larak r püskülünden elde edilen polifenoloksidaz aktivitesinin s cakl kla de is imi 20, 30, 40, 50, 60,70,80 ve 900 C’lerde incelenmi tir. Elde edilen verilerek Çizelge 3.7 ve Çizelge 3.8’de grafik edilmi tir. Katekol subsrat kullan ld nda m r püskülü PFO enziminin optimum s cakl 40O C, 4-metil katekol kullan ld nda ise optimum s cakl k 30ºC olarak tespit edildi. Literatür ara rmas nda ise Do an ve ark. (2005) enginar PFO enziminin optimum s cakl 40O C, Sun ve ark.(2010) chufa m n PFO enziminin optimum s cakl 45O C olarak tespit etmi lerdir.
Çizelge 3.7. M r püskülünden elde edilen polifenol oksidaz aktivitesinin katekol substrat nda s cakl k ile de imi
33
Çizelge 3.8. r püskülünden elde edilen polifenoloksidaz aktivitesinin 4-metilkatekol substrat nda s cakl k ile de imi
3.5. Termal inaktivasyon
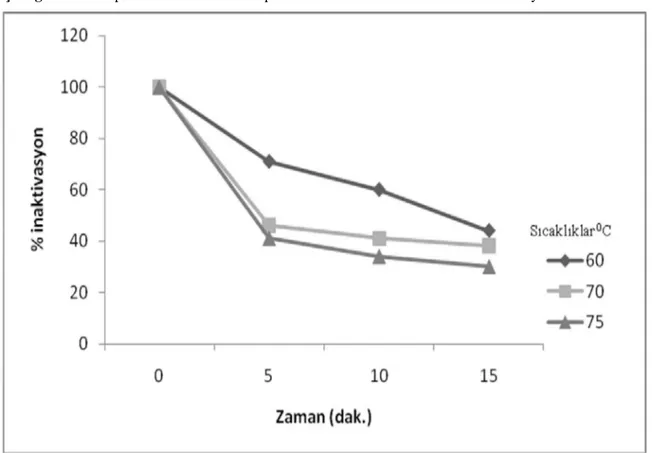
r püskülünden elde edilen polifenoloksidaz aktivitesinin termal denatürasyonu artan s cakl n ve inaktivasyon süresinin bir fonksiyonu olarak incelendi. Elde edilen deneysel veriler Çizelge 3.9’da grafik edildi. Deneysel bulgular tüm substratlar için artan s cakl k ve inaktivasyon süresi ile enzim aktivitesinin azald göstermektedir.
34
Çizelge 3.9. r püskülünden elde edilen polifenoloksidaz enziminin termal denatürasyon
3.6. Polifenoloksidaz aktivasyonu
Sukroz, fruktoz, riboz ve glukoz karbonhidratlar n m r püskülü PFO aktivitesi üzerine etkisi ara lm r. Fruktoz ve ribozun enzimi % 17 oran nda aktif etti i, glikozun % 16, sukrozun ise % 4 oran nda aktif etti i tespit edilmi tir. PFO enziminin aktivasyonu üzerine yap lan çal malar çok azd r. Bu durum ço u zaman, enzimin aktivasyonunun arzu edilmemesinden kaynaklanmaktad r. Çünkü kararma baz durumlar d nda (çay bitkisin kararmas ) istenmeyen bir durumdur. SDS’nin farkl avakado türlerinden haz rlanan ham ve
smen saflast lm ekstraktlardaki inaktif PFO’yu da aktive etti i bildirilmektedir. Ayr ca bakla PFO’sunun üre taraf ndan aktive edildi i de tespit edilmi tir (çizelge 3.10).
35
Çizelge 3.10. r püskülünden elde edilen polifenoloksidaz aktivitesine karbonhidratlar n etkisi
Karbonhidrat Konsantrasyon(k) Aktivasyon (%)
Glikoz 0.5 mM 2 mM 12 16 Sukroz 0.5 mM 2 mM 4 4 Fruktoz 0.5 mM 2 mM 4.5 17 Riboz 0.5 mM 2 mM 12.6 17 3.7. Polifenoloksidaz n inhibisyonu
EDTA,SDS ve sodyum azidin PFO aktivitesi üzerine inhibisyon etkisi katekol substrat kullan larak etkisi incelendi. Sonuçlar Çizelge 3.11’de gösterilmi tir. Bu çal mada sodyum azidin % 26.6 EDTA’n n % 22.2 ve SDS’nin % 12.2 oran nda enzimi etti i tespit edildi. nhibitör maddeler, reaksiyon ortam na eklendi inde enzim h azaltan do al ya da yapay kimyasal maddelerdir. PFO bir metallo proteindir. elat yap ajanlarla inhibe edilebilir ((EDTA= etilendiamintetraasetat), ve (sodyum azid)). (Zawistowski ve ark. 1991). Litreatürde marulun sodyum azid taraf ndan inhibe edilmedi i (Salman 2006) tespit edilmi tir.
Çizelge 3.11. r püskülünden elde edilen polifenol oksidaz aktivitesine inhibitörlerin etkisi
nhibitör Konsantrasyon( ) nhibisyon (%)
EDTA 0.5 mM 2 mM 16 mM 22.2 mM SDS 0.5 mM 2 mM 6.7 mM 12,2 mM Sodyum azid 0.5 mM 2 mM 11.1 mM 26.6 mM Nurten ASLAN
36 3.8. PFO’nun Homojenitesi ve Molekül A rl
Birçok kaynaktan elde edilen PFO enziminin farkl moleküler yap larda oldu u bildirilmektedir. Bu yap lar n say enzim kayna na ve enzimin ekstraksiyonunda ve saflas lmas nda uygulanan metotlara ba r. M r püskülü PFO enziminin moleküler
rl SDS-PAGE analizi sonucunda 158 kDa olrak belirlendi ( ekil 3.4). Egg plant tan elde edilen PFO enziminin moleküler a rl 112 kDa (Mishra et al. 2012), mango PFO enziminin moleküler a rl 31,5 kDa (Palma-Orozco et al. 2014), atemoya meyvesi PFO enziminin moleküler a rl ise 82 kDa olrak tespit edilmi tir (Chaves et al. 2011).
37 4. SONUÇ VE ÖNER LER
Bu çal mada m r püskülünden izole edilerek safla lan polifenol oksidaz n (PFO) biyokimyasal özellikleri ara lm r. Bu ba lamda, enzimin optimum pH ve s cakl , substrat spesifikli i ve termal inaktivasyonu, inhibitörlerin ve karbonhidratlar n etkisi incelenmi tir. Elde edilen bulgulardan; denenen substratlara enzimin ilgisinin yüksek oldu u oldu u ve enzim ilgisinin en yüksek oldu u substrat n katekol oldu u, termal stabilitesinin yüksek oldu u, sodyum azidin % 26.6 EDTA’n n % 22.2 ve SDS’nin % 12.2 oran nda enzimi etti i, ayr ca fruktoz ve ribozun enzimi % 17 oran nda aktif etti i, glikozun % 16, sukrozun ise % 4 oran nda aktif etti i ve enzimin moleküler a rl n 158 kDa oldu u tespit edilmi tir. Katekol subsrat kullan ld nda m r püskülü PFO enziminin optimum s cakl 40O C, 4-metil katekol kullan ld nda ise optimum s cakl k 30ºC olarak tespit edilmi , optimum pH de erleri ise katekol ve 4-metilkatekol substratlar kullan larak s ras yla 8-6 olarak tespit edilmi tir. Enzimatik kararman n önlenmesinde m r püskülünün pH’s n ve s cakl n bu aral klar n
nda tutulmas PFO enziminden kaynaklanan kararmay önemli ölçüde engelleyece i ifade edilebilir. Ekonomik de ere sahip m r bitkisi püskülü alternatif t pta da kullan ln maktad r. PFO enziminin ürünleri proteinlere ba lanarak söz konusu bitkinin besin de erini dü ürüp kokusunu ve tad de tirmektedir. Dolay yla bu çal ma m r püskülünün kullan lmas ve
lenmesi esnas ndaki kararman n önlenmesi aç ndan önemlidir.
Nurten ASLAN
39 5.KAYNAKLAR
Altunkaya, A. Gökmen, V.2013. Partial purification and characterization of polyphenoloxidase from durum wheat (Triticum durum L.). Journal of Cereal Science 55:300-304.
Arslan, O., Temur, A., ve Tozlu, . 1998. Polyphenol Oxidase from Malatya Apricot(Prunus armeniaca L.). J.Agr. Food Chem., 46:1239–1241.
Ayaz, F.A., Demir, O., Torun, H., Kolcuoglu, Y., ve Colak, A. 2008. Charecterization of polyphenoloxidase (PPO) and total phenolic contents in medlar (Mespilus germenica L.) fruit during ripening and over ripening. Food Chem., 106: 291–298.
Aydemir, T. 2004. Partial purification and charecterization of polyphenol oxidase from artichoke (Cynara scolymus L.) heads. Food Chem., 87: 59–67.
Chaves, I.R., De Souza Ferreira, E., Da Silva, M.A., Neves, V.A.2011. Polyphenoloxidase from atemoya fruit (Annona cherimola mill. Annona squamosa L.). J. Food Biochem. 35: 1583–1592.
Cheema S., Sommerhalter M.2015. Characterization of polyphenol oxidase activity in Ataulfo mango. Food Chemistry, 171: 382–387.
Constabel C.P., Barbehenn R., 2008.Induced Plant Resistance to Herbivory, Springer, Dordrecht , 253-269.
Corsini, D. L., Pavek, J. J. and Deab, B. 1992. Difference in free and protein-bound tyrosine among potate genotypes and their relationship to internal balackspot resistance. Am. Pot. J. 69: 423-434.
Dogan, M., Arslan, O., Dogan ,S. 2002. Substrat specifity, heat inactivation and inhibition of polyphenol oxidase from different aubergine cultivars. International Journal of Food Science and Technology, 37: 415–423.
Do an, S. 2002. Orignum L. (Lamiaceae) Taksonlar n (Origanum onites L. Ve Origanum vulgare L. spp.hirtum (Link) letswaar) Çevre Faktörleriyle Olan li kilerinin ve PFO Aktivitesinin Belirlenmesi. Doktora Tezi, Bal kesir Üniversitesi Fen Bilimleri Enstitüsü, Bal kesir.
Do an, S., Do an, M., Arslan, O. 2003. Characterization of polyphenol oxidase from Thymus(Thymus longicaulis var.subisophyllus).Advances in Food Science, 25(2):56-64.
Do an, S., Turan, P., Do an, M., Alkan, M.2006. Inhibition of Polyphenol Oxidase by Glutamic Acid. Process Biochemisrty. 41: 2379–2385.