İmatinib dirençli K562R hücre hattında kemoterapötik ajanların oluşturduğu reaktif oksijen türlerinin sığla yağı ile ortadan kaldırılması
Tam metin
Şekil
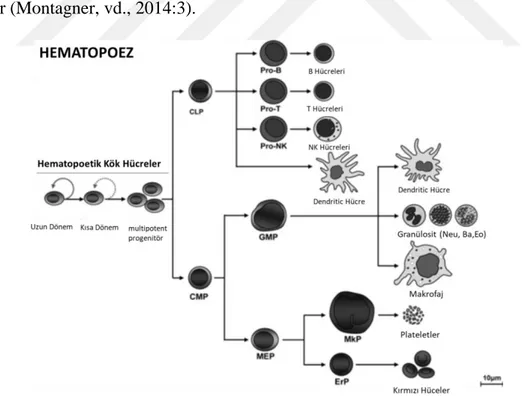
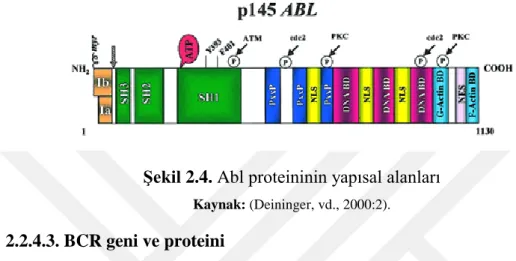
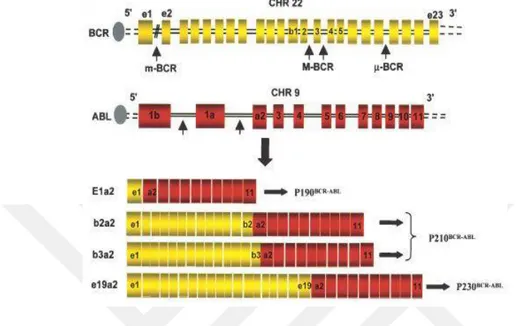
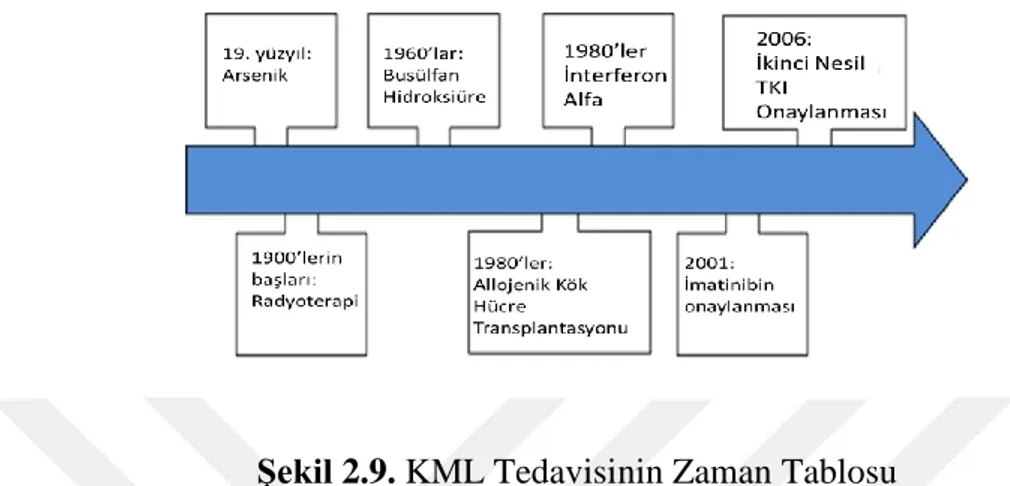

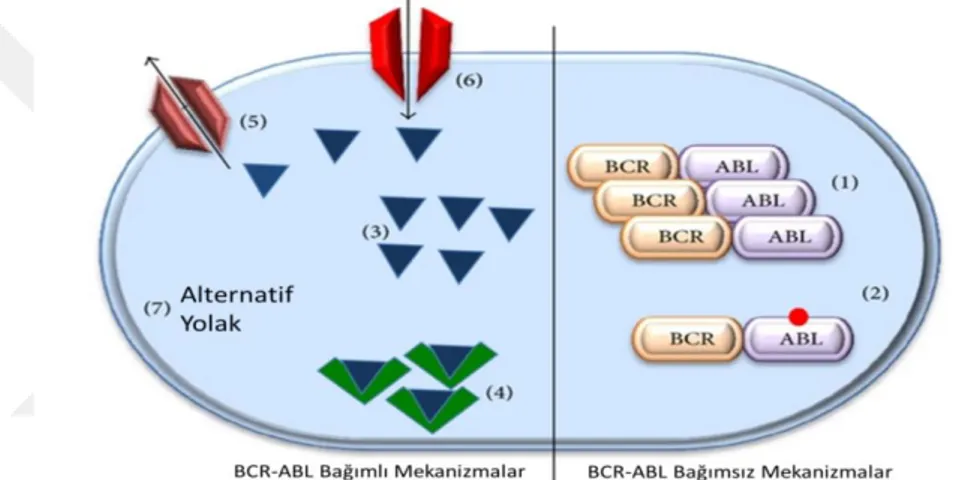
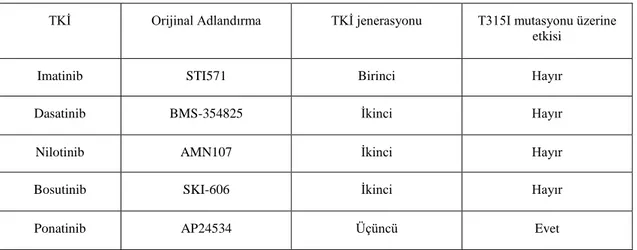

Benzer Belgeler
Doğa Koruma Merkezi, sığla ormanlarının devamlılığı için koruma biyolojisi ilkeleri ve peyzaj ekolojisi temel yaklaşımı ile bir koruma stratejisi öneriyor:
Republic of Iran, Iraq, Kuwait, Saudi Arabia and Venezuela.. They were to become the Founder Members of
Cupri oksit, bakır glukonat, bakır sulfat ve bakır aminoasit şelat halinde
Bilindiği gibi; basit filtrasyondan geçmiş ana faz (süt) içindeki, filtrelerin ayıramadığı katı, yarı katı veya yarı sıvı fazların santrifüj kuvveti ile
Asidik doğası gereği patojenler 24 saatte büyük ölçüde inaktive olmaktadır. Maya kontaminasyonu (Kluyveromyces and
ÇalıĢmamızda histopatolojik değerlendirmelerde yara iyileĢmesinin aĢamaları olan inflamasyon, kollajen birikimi, anjiogenezis, granülasyon dokusunun oluĢumu ve
Öz: Bu çalışmada, atık kızartma yağından elde edilen biyodizel, tek silindirli direkt enjeksiyonlu bir motorda yakıt olarak kullanılmış, performans, yanma ve emisyon
Bu çalışma kağıt fabrikalarında atık olarak ortaya çıkan tall yağından, yine şeker fabrikalarında atık statüsünde olan fuzel yağının alkol olarak kullanılması
