Mesenchymal stem cells and ligand incorporation
in biomimetic poly(ethylene glycol) hydrogels
signi
ficantly improve insulin secretion from
pancreatic islets
Tuğba Bal
1, Caner Nazli
2, Alparslan Okcu
3, Gökhan Duruksu
3, Erdal Karaöz
3,4and Seda Kizilel
1,2*
1Chemical and Biological Engineering, Koc University, Istanbul, Turkey
2
Material Sciences and Engineering, Koc University, Istanbul, Turkey
3
Center for Stem Cell and Gene Therapies Research and Practice, Kocaeli University, Kocaeli, Turkey
4
Liv Hospital, Regenerative Medicine, Stem Cell Research and Therapy Center, Istanbul, Turkey
Abstract
The main goal of this study was to investigate pancreatic islet function with mesenchymal stem cells (MSCs) in a ligand-functionalized poly(ethylene glycol) (PEG) hydrogel for the treatment of type 1 diabe-tes (T1D). Rat bone marrow-derived MSCs (rBM-MSCs) were encapsulated within synthetic PEG hydro-gel, and cell viability and apoptosis within this 3D environment was examined in detail. ATP content and caspase-3 activity of encapsulated MSCs showed thatfibronectin-derived RGDS, laminin-derived IKVAV and/or insulinotropic glucagon-like peptide (GLP-1) were required to maintain MSC survival. In-corporation of these peptides into the hydrogel environment also improved pancreatic islet viability, where combinations of peptides had altered effects on islet survival. GLP-1 alone was the leading stimu-lator for insulin secretion. Cell adhesion peptides RGDS and IKVAV improved insulin secretion only when they were used in combination, but could not surpass the effect of GLP-1. Further, when pancreatic islets were co-encapsulated with MSCs within synthetic PEG hydrogel, a two-fold increase in the stimulation index was measured. Synergistic effects of MSCs and peptides were observed, with a seven-fold increase in the stimulation index. The results are promising and suggest that simultaneous incorporation of MSCs and ECM-derived peptides and/or GLP-1 can improve pancreatic islet function in response to altered glucose levels in the physiological environment. Copyright © 2014 John Wiley & Sons, Ltd.
Received 28 November 2013; Revised 7 July 2014; Accepted 25 September 2014
Keywords islets; mesenchymal stem cells; PEG hydrogel; ligand incorporation; IKVAV; RGDS; GLP-1; visible light photopolymerization
1. Introduction
Type 1 diabetes (T1D) is an autoimmune disease and oc-curs as a result of pancreaticβ cell destruction, mediated by T cells as key players. These events lead to depletion of insulin, which drives hyperglycaemia and causes other severe problems, mostly at the vascular level (Diana et al., 2011; Kizilel et al., 2005). These complications in-clude increased function of endothelin-1 (ET-1), upregu-lation of endothelial nitric oxide synthase (eNOS) and
generation of more reactive oxygen species (ROS), resulting in blood flow and vessel abnormalities, and DNA damage (Brownlee, 2005; Diana et al., 2011). Clini-cal treatment of T1D requires daily control of blood glu-cose levels and several doses of daily insulin injections. Alternatively, oral/nasal insulin delivery and islet trans-plantation are used in clinical practice to maintain euglycaemia. However, shortage of donor tissue and the requirement for the use of immunosuppressive drugs are some of the main limitations for clinical application of islet transplantation (Lin and Anseth, 2009; Matsumoto, 2010). To date, various strategies have been developed to eliminate immune suppression, where conformal coatings with living cells (e.g. Treg, HEK293) (Marek et al., 2011;
*Correspondence to: Seda Kizilel, Chemical and Biological Engineering, Koc University, Istanbul 34450, Turkey. E-mail: [email protected] J Tissue Eng Regen Med 2017; 11: 694–703.
Teramura and Iwata, 2010), molecule immobilization (e.g. urokinase) with layer-by-layer assembly on islet surfaces (Kizilel et al., 2010; Teramura and Iwata, 2010; Teramura et al., 2013) or encapsulation of pancreatic islets within hy-drogel capsules, e.g. poly(ethylene glycol) (PEG), colla-gen, alginate, have been considered (Liao et al., 2013; Lin and Anseth, 2009; Lin et al., 2010; O’Sullivan et al., 2011; Weber and Anseth, 2008). These previous efforts were successful to a limited extent, and further investiga-tion is required to achieve long-term islet funcinvestiga-tion in clinical transplantation studies. Mesenchymal stem cells (MSCs) have been found to enhance islet graft vascularization and viability, reverse diabetes (Ito et al., 2010; Sims and Evans-Molina, 2012) and reduce blood coagulation (com-pared to controls) and apoptosis when cultured in vitro or implanted with pancreatic islets (Duprez et al., 2011; Yeung et al., 2012). Recently, Kerby et al. (2013) and Davis et al. (2012) reported positive influences of MSCs on pancreatic islets within alginate microcapsule or silk hydrogel macrocapsule environment, where an approximately two-fold increase in insulin secretion could be obtained from en-capsulated islets compared to control groups.
In the present study, we studied the function of encap-sulated pancreatic islets and the anti-apoptotic properties of MSCs in ligand-functionalized PEG hydrogel scaffolds. The synthetic hydrogel matrix includes adhesion peptides (RGDS and IKVAV) and an insulinotropic peptide (GLP-1) in combination with rat bone marrow-derived MSCs (rBM-MSCs). In the system considered here, PEG hydro-gel membrane acts not only as a mechanical barrier but also as a functional synthetic extracellular matrix (ECM), with covalently bonded biomolecules for the maintenance of islets and MSCs in a three-dimensional (3D) network. We observed that MSCs supported encapsulated islets within the PEG hydrogel environment and increased insu-lin secretion upon elevated concentrations of glucose exposure. Finally, acrylated RGDS and IKVAV contributed to the viability of the MSCs during 5 days of in vitro incuba-tion, and MSCs further supported the function of pancreatic islets within the PEG hydrogels. The results presented here suggest that MSCs, in combination with ECM-derived peptides and GLP-1, can promote the highest glucose-stimulated insulin secretion (GSIS) from pancreatic islets compared to the other systems considered in the literature, and are ideal candidates for the clinical reversal of T1D.
2. Materials and methods
2.1. Materials
Penicillin–streptomycin, histopaque,L-glutamine, eosin Y and 1-vinyl-2-pyrrolidone (NVP) were purchased from Sigma. L-DMEM, RPMI-1640 and Alamar blue were sup-plied from Invitrogen. Fetal bovine serum (FBS; heat-inactivated), triethanolamine (TEA) and collagenase P were from Biological Industries, Merck and Roche, respec-tively. Poly(ethylene glycol) diacrylate (PEGDA) was the
product of Laysan Bio Inc. All tissue-culture plates were from Costar, unless otherwise stated. CellTiter-Glo®
Luminescent Cell Viability Assay and Caspase Glo 3/7 Reagent were purchased from Promega (Madison, WI, USA). Rat insulin ELISA was obtained from Mercodia AB.
2.2. rBM-MSC isolation, culture and
encapsulation
rBM-MSCs isolation was performed as described previously (Karaoz et al., 2009). Following isolation and characteriza-tion, MSCs were cultured in L-DMEM supplemented with 10% FBS and 100 IU/ml penicillin–0.1 mg/ml streptomycin at 37°C in a 5% CO2environment. The MSCs were passaged
every 3–4 days. rBM-MSCs were suspended in the prepolymer solution, consisting of 10% w/v PEGDA (10 kDa), 225 mM
TEA, 37 mMNVP and 0.03 mMeosin Y. For specific sample sets, 2.5 mMacrylate-PEG-RGDS (acr-PEG-RGDS), 2.5 mM
acr-PEG–IKVAV and/or 20 μM acr-PEG–GLP-1 were in-cluded in the solution; 30μl of this prepolymer solution, with 20 000 cells, was photopolymerized at 50 mW (514 nm, argon ion laser; Coherent Inc., Santa Clara, CA, USA) for 3 min. After photopolymerization, the encapsu-lated cells were placed into 24-well plates and incubated with the growth medium at 37°C under 5% CO2. The cell
culture medium was changed every other day.
2.3. Pancreatic islet isolation and encapsulation
Pancreatic islets were isolated from Wistar rats by enlarg-ing the pancreatic duct with a solution of collagenase P. Following digestion at 37°C, a histopaque density gradient was applied to purify the islets. After handpicking, islets were transferred to RPMI-1640 medium supplemented with 10% FBS, 2 mM L-glutamine and 100 IU/ml penicillin– 0.1 mg/ml streptomycin at 37°C in a 5% CO2environment
and were cultured overnight (Omer et al., 2004).
For encapsulation experiments, ~20 islets were suspended in 30μl prepolymer solution with or without peptide con-jugates, and were exposed to a laser at 50 mW for 3 min. The hydrogel samples were transferred to 24-well plates and cultured for 3 days.
2.4. Co-encapsulation of islets and rBM-MSCs
Twenty islets were suspended in 30μl PEGDA prepolymer solution with 20 000 MSCs. Next, this solution was ex-posed to green light at 50 mW for 3 min (Scheme 1). En-capsulated MSCs and islets in hydrogels were cultured for 3 days.
2.5. Viability and caspase activity
The viability of the encapsulated cells was assessed with CellTiter-Glo® Luminescent Cell Viability Assay. Briefly, cell-containing hydrogels were incubated with 50%
Cell-Titer Glo reagent for 1.5 h at room temperature. ATP standards were used to generate a standard curve. The resulting luminescence was measured using a microplate reader (Synergy H1, Bio-Tek). The measured ATP amounts were normalized with Alamar blue reagent for the specific hydrogel sample, to prevent differences in cell numbers for islet groups. Briefly, hydrogels were first treated with 10% Alamar blue reagent for 3 h and then the measured fluorescence signal (Ex: 550 nm, Em: 590 nm) was used to monitor viable cells (Lin and Anseth, 2009).
For the caspase activity, encapsulated cells were incu-bated with 50% Caspase Glo 3/7 Reagent for 1.5 h and luminescence was measured. In order to prevent variation in cell numbers between different groups, normalization with respect to Alamar bluefluorescence was carried out for the specific hydrogel sample to calculate apoptotic activity.
2.6. Static incubation assay
The functionality of encapsulated/co-encapsulated islets was tested via a static incubation assay. Encapsulated islets were placed into low (2.8 mM) and high (28 mM) glucose buffers for 1 h. After each incubation period, sam-ples were collected and analysed with rat insulin ELISA to measure insulin content. Insulin concentration in low-and high-glucose buffers were then used to calculate the stimulation index.
2.7. Statistical analysis
All datasets were expressed as mean ± SD. Statistical analysis was performed with one-way ANOVA with Bonferroni’s test; p < 0.05 was considered significant.
3. Results
3.1. Viability and caspase activity of rBM-MSCs
in PEG hydrogels
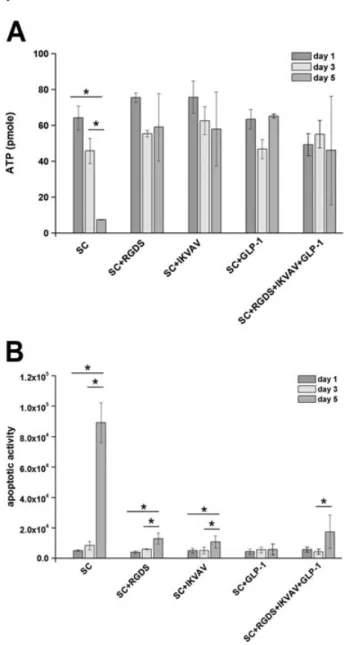
Encapsulation of MSCs within cell adhesion peptides and GLP-1-functionalized, covalently crosslinked PEG hydrogels was carried out via visible light photopolymerization. To evaluate the morphology of MSCs upon encapsulation, cells were observed for 5 days. For the conditions studied, the morphology of MSCs remained spherical in PEG hydrogels and no significant changes were observed until day 5 (Figure 1). In contrast, altered survival rates were mea-sured for MSCs encapsulated within functional PEG hydro-gel. In the absence of ligands, decreasing ATP values were measured for days 3 and 5. This was attributed to reduced metabolic activity, which showed that during this incuba-tion period, the viability of the cells was decreased to 72.0 ± 11.0% on day 3 and 11 ± 0.01% on day 5, compared to the viability measured on day 1 (Figure 2A). In the pres-ence of peptide conjugates, slight decreases in cell viabil-ities were observed, where the lowest ATP measured was around 50 pMon day 5. When RGDS, IKVAV, GLP-1 or
com-bination of these peptides was covalently bonded to the sur-rounding hydrogel network, the ATP values of MSCs were higher than those of cells encapsulated in unmodified hydrogels.
The effect of ligand incorporation on MSC viability was also supported by an apoptosis assay, as apoptotic activi-ties measured for unmodified hydrogels and peptide-immobilized hydrogels were similar on day 1 (Figure 2B). When the MSCs were encapsulated within unmodified hydrogels, apoptotic activity increased from 5001 ± 492 on day 1 to 8357 ± 2751 on day 3 and 89142 ± 13125 on day 5. As represented in Figure 2B, longer incubation
Scheme 1. Encapsulation of islets and rBM-MSCs in ligand-functionalized PEG hydrogel scaffold. [Colourfigure can be viewed at wileyonlinelibrary.com]
periods resulted in slight increases in caspase activities when single or combinations of peptides were bonded to PEG hydrogel scaffold. Interestingly, positive influences of peptides on the apoptosis of encapsulated MSCs could only be observed on day 5, when significantly lower values of apoptotic activity were measured compared to the control group (unmodified hydrogels).
3.2. Effects of ligand incorporation on islet
viability and caspase activity
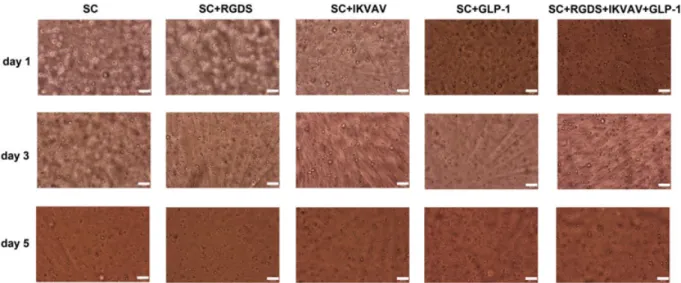
The influence of peptides and MSCs incorporation within a PEG hydrogel network was assessed through measure-ment of ATP content and caspase activity for islet survival on day 3. Figure 3 illustrates optical microscope images of islets encapsulated within PEG hydrogel scaffolds for var-ious conditions, including ligands and/or MSCs. It was ob-served that encapsulated islet morphology was similar in the presence or absence of MSCs, with altered conditions of ligand functionalization.
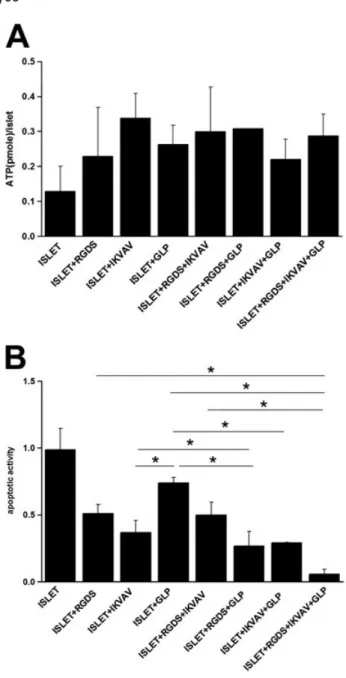
Encapsulated islets within PEG hydrogels resulted in similar ATP values (Figure 4A). The amount of ATP/islet was around 0.13 pM, which has similar order of
magni-tude to, and is consistent with, the previous literature studies about pancreatic islets in biomimetic matrices (Lin and Anseth, 2009). ATP values measured for islets encapsulated in ligand-conjugated hydrogels increased to 3.2 ± 1.6-fold of those of the islets encapsulated in un-modified hydrogel control group.
Caspase activity for islets encapsulated in unmodified hydrogels was significantly higher than in the other re-maining groups, where apoptotic activity of 0.99 ± 0.16 was obtained for this control group (Figure 4B). Apoptotic activities of islets encapsulated in the presence of cova-lently attached peptides were 0.51 ± 0.07 for RGDS-, 0.37 ± 0.09 for IKVAV- and 0.74 ± 0.04 for GLP-1-containing groups. Simultaneous incorporation of peptides
decreased apoptotic activity to at most 0.27 ± 0.11-fold of the control group. Caspase activity, and hence apoptotic index, was further reduced to 0.06 ± 0.03 with respect to the control group when all three peptides were conjugated to PEG hydrogel scaffolds (Figure 4B).
3.3. Viability of co-encapsulated islets and
rBM-MSCs in PEG hHydrogels
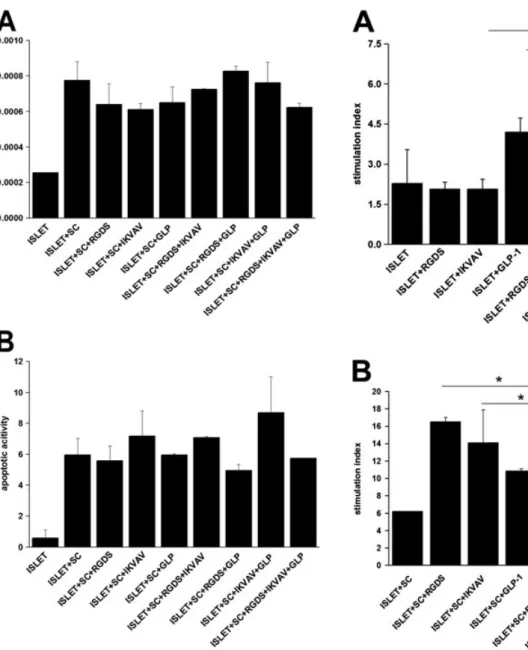
Intracellular ATP amount and caspase activities were also measured to examine the effects of peptide incorporation on the total viability of co-encapsulated cells. ATP concen-trations are shown in Figure 5A, where the total viability of co-encapsulated cells was in the range 0.8–1.1. Al-though combinations of ligands increased ATP levels slightly, these effects were estimated to be insignificant. ATP results were also supported by caspase activity assay. Apoptotic activities were within the range 5.7–8.6 (Fig-ure 5B). This suggests that the numbers of viable cells for different encapsulation groups were comparable.
3.4. Functionality of encapsulated islets in the
presence of covalently immobilized ligands
and/or rBM-MSCs
For the functionality assay, encapsulated islets were ex-posed to glucose challenge with a static incubation assay, and the capability of islets to secrete insulin in response to altered glucose concentrations was expressed as the stim-ulation index (Figure 6). The stimstim-ulation index represents the ratio of the amount of insulin secreted by islets at high glucose concentration to the amount of insulin secreted at low glucose concentration. This gives more accurate infor-mation about GSIS from islets as a clinical release crite-rion, as in each set with similar islet numbers, the number of cells within each islet might differ, which could
Figure 1. Optical microscope images of rBM-MSCs encapsulated in biofunctional PEG hydrogels. MSCs were encapsulated (20 000 cells/hydrogel) and cultured in DMEM containing 10% FBS and 1% penicillin–streptomycin. Visualization with light microscopy was performed on days 1, 3 and 5 post-encapsulation; hydrogel volume = 30μl; scale bars = 100 μm. [Colour figure can be viewed at wileyonlinelibrary.com]
be misleading when expressed as insulin amount at low and high glucose concentrations (Mueller et al., 2013). This was also used in previous recent studies about encap-sulated islets to characterize insulin secretion function (Davis et al., 2012; Lin and Anseth, 2011). Thus, in each case, change of islet responsiveness against glucose levels under studied conditions would clearly show the role of peptides or MSCs. The stimulation indices of islets in un-modified, RGDS- and IKVAV-modified PEG hydrogels were approximately similar at ~2. Combinations of RGDS and IKVAV induced islet functionality further by increasing the stimulation index to 4.92 ± 0.13. The insulinotropic peptide GLP-1 was shown to be more effective than ECM
peptides (Figure 6A). The addition of IKVAV, along with GLP-1, into the encapsulation environment promoted islet function further, and this suggests that the addition IKVAV and GLP-1 has a synergistic effect to promote insulin se-cretion in response to high glucose. Further incorporation of RGDS into IKVAV + GLP-1 hydrogel was also a trigger for islet function, and a similar stimulation index value of 4.57 ± 2.19 was measured for the islets encapsulated in this group.
The capacity of islets to respond to altered levels of glu-cose was improved significantly in the presence of MSCs (Figure 6B). Co-encapsulation of islets and MSCs in unmodified PEG hydrogel scaffolds resulted in a two-fold increase in the stimulation index compared to the islet-only condition. We observed substantial improvements when MSCs and islets were encapsulated within ligand-incorporating PEG hydrogel scaffolds. The stimulation in-dex further rose to seven-fold of the islet-only group when peptides and MSCs were used in combination.
4. Discussion
A bioartificial pancreas in a functional synthetic hydrogel scaffold would be influenced by the signals within the synthetic environment. The viability and functionality of islets are dictated by these signals, and the effects of li-gand incorporation on islet function and viability should be investigated in detail before a bioartificial pancreas can become a clinical reality. Here, we report that pancre-atic islet viability and functionality in 3D networks such as PEG hydrogels can be improved through co-encapsulation with MSCs within ligand-incorporating PEG hydrogel scaffolds. The reason for covalent incorporation of pep-tides was to enhance their physiological effect on islet sur-vival. For example, insulinotropic ligand GLP-1, used here, would be cleaved by dipeptidyl-peptidase within nearly 2 min (Li et al., 2011) when administered nously. However, in our system there is no need for intrave-nous administration of GLP-1, as its covalent incorporation would keep the ligand stationary within the synthetic network and would not involve plasma circulation, when considered for implantation into an in vivo model. As shown here, covalent incorporation of peptides increased viability to 320%, and the stimulation index was enhanced by two-fold compared to control islets. Further, significant enhancement in islet function was observed with co-encapsulation of islets with rBM-MSCs. A 3-day culture period was chosen for the duration of co-encapsulation experiments, as MSCs would eventually differentiate into adipogenic, osteogenic or chondrogenic lineages (Anderson et al., 2011; Muraglia et al., 2000). The 3-day in vitro culture experiments proved that MSCs could still induce insulin secretion from islets. Further, in unmodified hydrogels, MSCs survival would be significantly compromised on day 5 and no stimulatory effect would be expected.
In the presence of MSCs and ligands, the functionality of islets could be increased up to seven-fold compared to islets encapsulated alone. These results support the idea
Figure 2. Effect of covalent immobilization of peptides in PEG hydrogels on rBM-MSCs survival. The MSCs were encapsulated (20 000/hydrogel) and maintained in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% FBS and 1% penicillin– streptomycin. (A) At specified time points, the amount of intracellu-lar ATP was measured as an indicator of MSC viability (mean ± SD). (B) Caspase activity was assessed on days 1, 3 and 5 to generate cel-lular apoptotic activity data. MSC-laden hydrogels were exposed to caspase activity reagent and luminescence was normalized to the amount of ATP measured under the same conditions for apoptotic activity (mean ± SD); hydrogel volume = 30μl; *p < 0.05.
of enhanced insulin release metabolism of encapsulated islets with an appropriate cell type in the presence of ECM-derived biofunctional signalling peptides and GLP-1. The viability and caspase activity of rBM-MSCs were ex-amined through encapsulation of cells within cell adhe-sion ligand- and GLP-1-incorporating PEGDA scaffolds.
The results demonstrated the dependence of MSC via-bility on cell adhesion peptides and GLP-1 (Figure 2). In the absence of adhesion peptides, due to the lack of cell–matrix signalling-related cell attachment, cells begin to increase caspase activity and enter apoptosis, mainly anoikis, which occurs as a result of detaching from the surrounding ECM (Weber et al., 2007). This results in massive cell death in in vitro cell culture. Our observa-tions demonstrated that 90% of cells lost viability on day 5, which is also consistent with previousfindings. For ex-ample, Nuttelman et al. (2005) also reported that 85% of MSCs were not viable in unmodified PEG hydrogel scaf-folds after 7 days of incubation.
Encapsulation of MSCs in the presence of cell adhesion ligands and insulinotropic ligands within the PEG hydro-gel scaffolds significantly improved MSCs survival (Fig-ure 2). ATP amounts and caspase activities meas(Fig-ured for each encapsulation condition illustrated that, despite slight differences in intracellular ATP amount and caspase activity, MSCs were still metabolically active and>85% of cells were viable regardless of the type of peptide ligand used. This is due to the role of RGDS and IKVAV to support various processes, including adhesion, proliferation and spreading through interaction with integrins (Hersel et al., 2003; Sreejalekshmi and Nair, 2011; Weber et al., 2007). Lee et al. (2010, 2012) have also reported that the RGDS- or IKVAV-binding integrins α5β1, αvβ5 and
α6β1 are crucial for the expression of stemness-related
genes within PEG hydrogel scaffolds for at least 5 days. This property is important for stem cell potency to
promote GSIS from islets within synthetic 3D encapsula-tion systems. In the long term, these physically entrapped MSCs within PEG hydrogels may continue to migrate, and proliferate if the hydrogel is degradable. The PEG– hydrogel system applied here is covalently crosslinked and non-degradable, with an average pore size of around 10 nm, which is smaller than the diameter of MSCs (~10μm). As a result, no significant migration and growth will occur (Anderson et al., 2011; Chan et al., 2007). How-ever, the duration of multipotency, the risk of a role in tumor formation or directed differentiation to a desired cell lineage to support graft function in vivo needs to be investi-gated further, as recent studies showed that MSCs do not form tumors but can act as pericytes and, thus, increase the angiogenesis and metastasis of pre-existing tumor cells (Bexell et al., 2009; Zhu et al., 2006).
Interestingly, we observed that when MSCs were encap-sulated in GLP-1-functionalized PEG hydrogel scaffolds, viability and caspase activities were similar to RGDS- or IKVAV-conjugated conditions (Figure 2). To date, there is no report on the effect of GLP-1 on MSCs survival within synthetic networks, and it was demonstrated here for the first time that insulinotropic ligands such as GLP-1 played an important role in the survival of MSCs within the PEG hydrogel. Thisfinding here is consistent with those of the previous study in which Sanz et al. (2010) observed that GLP-1 prevented the differentiation of MSCs by interfering with the expression of PPARγ, C/EBP and LPL and in-creased cell proliferation and prevented apoptosis via the MEK and PKC pathways in an in vitro cell culture model.
The influence of biofunctional ligands within PEG hy-drogel scaffolds on pancreatic islet cell viability and apo-ptosis was also investigated in this study. The ATP concentrations and apoptotic activities of islets within the PEG hydrogel scaffolds differed from each other, and combinations of peptides did not provide additional
Figure 3. Optical microcope images of islets (20 islets/hydrogel) or islets + MSCs (20 islets + 20 000 MSCs/hydrogel) encapsulated within ligand functional PEG hydrogel scaffold ([IKVAV] = 2.5 mMand [GLP-1] = 20μM). Visualization was performed on day 3 post-encapsulation. The hydrogels were maintained in DMEM supplemented with 10% FBS and 1% penicillin–streptomycin. Encapsulated islets: (A) within unmodified PEG hydrogel; (B) within IKVAV functionalized hydrogel; (C) within GLP-1 functionalized hydrogel; (D) within IKVAV+GLP-1-functionalized PEG hydrogel; (E) with focusing on MSCs; and (F) with focusing on islets and rBM-MSCs in IKVAV+GLP-1-functionalized PEG hydrogel; black and red arrows, islets and rBM-MSCs, respectively; hydrogel volume = 30μl; scale bars = 50μm. [Colour figure can be viewed at wileyonlinelibrary.com]
benefits compared to the conditions in which individual peptides were used (Figure 4). The presence of GLP-1 or IKVAV was more effective than that of RGDS. This could be due to the interaction of the ligands used here with their specific receptors, such as the 110 kDa laminin recep-tor that is abundant in the basement membrane of islets for IKVAV (Nikolova et al., 2006; Weber et al., 2007, 2008) and the G-protein-coupled receptor (GPCR) of GLP-1,
activating various signalling pathways, such as the focal ad-hesion kinase phosphorylation, protein kinase A (PKA) and phosphoinositide-3 (PI-3) kinase pathways (Doyle and Egan, 2007; Drucker, 2001; Vahl et al., 2003). Such specific signalling pathways impose remarkable influences on the survival and function ofβ-cells encapsulated within PEG hy-drogel scaffolds (Lin and Anseth, 2009; Weber et al., 2007). In the absence of any ligand incorporation within the net-work, ATP per islet in unmodified hydrogels was lower, and apoptotic activities for islets were greater, than the values observed for islets within biofunctional PEG hydro-gel scaffolds. This was expected, as it was well established in prior studies that the PEG hydrogel network acts like a blank slate for encapsulated cells (Lin and Anseth, 2009).
To further investigate anti-apoptotic properties in the presence of MSCs, islets were co-encapsulated with MSCs in the presence or absence of covalently bonded ligands within PEG hydrogel scaffolds. The presentation of ECM mimic peptides (RGDS and IKVAV) reduced islet apoptosis (Figure 4B), while with co-encapsulated MSCs, islet apoptosis was not significantly different in the presence of ECM peptides compared to control (islets + MSCs) (Figure 5B). This could be due to the significant apoptosis of MSCs rather than islets. Also, according to Figure 5A, the amount of ATP was not different in co-encapsulated islets and MSCs, with or without presentation of peptides in the scaffold. This might indicate that the majority of the signals were coming from MSCs, and that the presenta-tion of peptides in hydrogel scaffolds did not have a pro-found effect on apoptosis of the co-encapsulated islets and MSCs. However, several prior studies reported the anti-apoptotic effect of MSCs on islets in cell culture me-dium, and this was not observed here (Figure 5). This was most probably due to differences in the experimental approaches used here and in other studies. Previous stud-ies focused mainly on reducing the harmful effects of cyto-kines [tumour necrosis factor-α (TNFα), interleukin-1β (IL-1β), interferon-γ (IFNγ)] or streptozotocin (STZ) and hypoxia-induced apoptosis of islets in a co-culture with MSCs in vivo and in vitro (Abdi et al., 2008; Bhaiji et al., 2012; Davis et al., 2012; Duprez et al., 2011; Karaoz et al., 2010; Lu et al., 2010; Yeung et al., 2012). Here, we mainly investigated the protection potential of islets from apoptosis with the anti-apoptotic property of MSCs in the presence or absence of biofunctional signalling molecules (Frisch and Screaton, 2001; Grossmann, 2002). Presenta-tion of ligands within 3D PEG hydrogel scaffolds for islets and MSCs provided sufficient cell–matrix interactions; how-ever, cell–cell interaction is also required to prevent apopto-tic pathways. In the system considered here, investigation of the influence of cell–cell signalling pathways on islet via-bility or function was not within the scope of this study and was not considered. As a result, viabilities and caspase activ-ities were observed to be similar and the anti-apoptotic properties of MSCs on islets could not be observed.
It is also noteworthy that the ligands had a positive in-fluence on islet function. RGDS- or IKVAV-incorporating scaffolds resulted in similar insulin secretion from islets, with a stimulation index value of ~2, where covalent
Figure 4. Effects of ECM-derived peptides on encapsulated islet viability. Islets were encapsulated in the presence of acr-PEG– RGDS (2.5 mM), acr-PEG–IKVAV (2.5 mM) and/or acr-PEG–GLP-1 (20μM). ATP amount, mitochondrial activity and caspase amounts were measured on day 3 post-encapsulation. (A) Intra-cellular ATP (pM; mean ± SD) and (B) caspase activity of encap-sulated islets within functionalized PEG hydrogels was assessed to compare apoptotic activities. Caspase luminescence of each condition was normalized with Alamar bluefluorescence of the specific sample. Further normalization was performed with num-ber of islets in the hydrogel for ATP amount, and with islet-only control groups for apoptotic activity (mean ± SD); *p < 0.05. Apoptotic activities of islets with GLP-1 and RGDS + IKVAV + GLP-1 differ statistically from all other conditions; hydrogel volume = 30μl
conjugation of GLP-1 into the matrix resulted in higher stimulation index values. This showed that GLP-1 alone was sufficient to promote GSIS, and cell-adhesion ligands had significant synergistic effects on insulin secretion by islets (Lin and Anseth, 2009; Weber and Anseth, 2008; Weber et al., 2007). These results, in combination with the viability assay, showed that adhesion peptides could support cell survival, whereas for the insulin secretion function of islets, GLP-1 should be present within the 3D
PEG hydrogel scaffold. IKVAV was superior to RGDS, which could be due to its interaction with the 110 kDa laminin receptor (Weber et al., 2007, 2008). This interac-tion would induce cytoskeletal alterainterac-tions related to F-actin and t-SNARE communication, as well as activation of nu-clear factor-κB (NF-κB) (Weber et al., 2007). These events
Figure 5. Effects of combination of rBM-MSCs and ligands on islet survival on day 3 post-encapsulation. Islets and MSCs (20 000/hydrogel) were co-encapsulated in the presence of acr-PEG–RGDS (2.5 mM), acr-PEG–IKVAV (2.5 mM) and/or acr-PEG– GLP-1 (20μM). (A) Total ATP amount (pM) of co-encapsulated islets and MSCs in PEG hydrogels were measured for cell viabil-ity. Encapsulated cells were first treated with Alamar blue for mitochondrial activity and then, the same samples were utilized for ATP measurements (mean ± SD). (B) Caspase activity of islets and rBM-MSCs in PEG hydrogels were represented as apoptotic activities. Similar to the normalization for ATP amount, samples werefirst used in Alamar blue assay, and in the following step, total caspase activity was measured for the specific hydrogel sample (mean ± SD); hydrogel volume = 30μl.
Figure 6. Effects of combination of rBM-MSCs and ligands on the functionality of islets encapsulated within PEG hydrogel on day 3 post-encapsulation. Islets or islets and MSCs (20 000/hydrogel) were encapsulated in the presence of acr-PEG–RGDS (2.5 mM), acr-PEG–IKVAV (2.5 mM) and/or acr-PEG–GLP-1 (20 μM). Cell-laden hydrogels were maintained in DMEM containing 10% FBS and 1% penicillin–streptomycin. Encapsulated cells were challenged with glucose assay on day 3. Encapsulated cells within PEG hydrogels were first treated with 2.8 and 28 mM glucose buffer for 2 h. The amount of insulin released was quantified with rat insulin ELISA. For the stimulation index, the amount of insulin released in 28 mMglucose buffer was normalized with amount of insulin released in 2.8 mMglucose buffer. (A) Stimulation index of islets encapsulated within peptide-modified PEG hydrogels (mean ± SD). (B) Stimulation index of co-encapsulated islets with MSCs in peptide-modified PEG hydrogels (mean ± SD); hydrogel volume = 30μl; *p < 0.05
would promote the synthesis of the proinsulin gene in β cells and the docking of insulin granules to the membrane, which are important for GSIS (Weber et al., 2007). GLP-1 is the most dominant stimulant compared to the other ligands used here, as this insulinotropic peptide acts onβ cell mass increase, insulin biosynthesis and release from β cells (Abraham et al., 2002; Doyle and Egan, 2007; Drucker, 2001; Mapelli et al., 2009; Vahl et al., 2003). Co-encapsulation of islets with MSCs also activated GSIS from islets at a similar level (Figure 6B). This could be explained as a result of the re-lease of trophic factors such as HGF and TGFβ from MSCs, rather than cell–cell contact-dependent islet–MSC signalling events, as direct contact between islets and MSCs was not designed (Figure 3) (Davis et al., 2012; Park et al., 2010). This also proves that tropic factor release from MSCs could promote GSIS; however, further modifications, such as coating of islets with MSCs, could also be considered before co-encapsulation within a 3D synthetic scaffold.
5. Conclusion
The functionality of islets can be significantly improved within 3D PEG hydrogels in the presence of rBM-MSCs, ECM-derived cell adhesion ligands and GLP-1. Significant improvements in islet functionality was observed on the co-encapsulation of islets with MSCs. Immobilized
peptides acted as ECM mimic ligands to stimulate cell sur-vival and insulin secretion pathways. GLP-1, a potent incretin hormone and a receptor agonist, promoted stimu-lation of insulin gene transcription, islet growth and neogenesis as a result of binding to its receptor on the surface of islets, and hence increased insulin secretion in response to high glucose. Further addition of MSCs into the system resulted in significant improvements in func-tionality, most probably due to soluble factor release from the MSCs. Synergistic effects of cell adhesion peptides on islet functionality were also observed. This study is prom-ising in demonstrating the combination of fundamental parts of a tissue (cell and ECM) and insulinotropic ligands for the future design of a bioartificial pancreas and for the treatment of T1D.
Con
flict of interest
The authors have declared that there is no conflict of interest.
Acknowledgements
We would like to acknowledge Istanbul Rotary Club and TUBITAK-MAG-113M232 for funding this project. The authors also thank Dr Nathan Lack, at Koc University School of Medicine, for providing help with luminescence andfluorescence measurements.
References
Abdi R, Fiorina P, Adra CN et al. 2008; Immunomodulation by mesenchymal stem cells: a potential therapeutic strat-egy for type 1 diabetes. Diabetes 57(7): 1759–1767.
Abraham EJ, Leech CA, Lin JC et al. 2002; Insulinotropic hormone glucagon-like peptide-1 differentiation of human pancre-atic islet-derived progenitor cells into insulin-producing cells. Endocrinology 143 (8): 3152–3161.
Anderson SB, Lin CC, Kuntzler DV et al. 2011; The performance of human mesenchymal stem cells encapsulated in cell-degradable polymer–peptide hydrogels. Biomaterials 32(14): 3564–3574.
Bexell D, Gunnarsson S, Tormin A et al. 2009; Bone marrow multipotent mesen-chymal stroma cells act as pericyte-like mi-gratory vehicles in experimental gliomas. Mol Ther 17(1): 183–190.
Bhaiji T, Zhi ZL, Pickup JC. 2012; Improving cellular function and immune protection via layer-by-layer nanocoating of pancre-atic isletβ-cell spheroids cocultured with mesenchymal stem cells. J Biomed Mater Res A 100(6): 1628–1636.
Brownlee M. 2005; The pathobiology of dia-betic complications: a unifying mecha-nism. Diabetes 54(6): 1615–1625. Chan BP, Hui TY, Yeung CW et al. 2007;
Self-assembled collagen-human mesenchymal stem cell microspheres for regenerative medicine. Biomaterials 28(31): 4652–4666.
Davis NE, Beenken-Rothkopf LN, Mirsoian A et al. 2012; Enhanced function of pancreatic islets co-encapsulated with ECM proteins and mesenchymal stromal cells in a silk hydrogel. Biomaterials 33(28): 6691–6697. Diana J, Gahzarian L, Simoni Y et al. 2011; Innate immunity in type 1 diabetes. Discov Med 11(61): 513–520.
Doyle ME, Egan JM. 2007; Mechanisms of action of glucagon-like peptide 1 in the pancreas. Pharmacol Ther 113(3): 546–593. Drucker DJ. 2001; Minireview: the glucagon-like peptides. Endocrinology 142(2): 521–527.
Duprez IR, Johansson U, Nilsson B et al. 2011; Preparatory studies of composite mesenchymal stem cell islets for applica-tion in intraportal islet transplantaapplica-tion. Ups J Med Sci 116(1): 8–17.
Frisch SM, Screaton RA. 2001; Anoikis mecha-nisms. Curr Opin Cell Biol 13(5): 555–562. Grossmann J. 2002; Molecular mechanisms of
‘detachment-induced apoptosis – anoikis’. Apoptosis 7(3): 247–260.
Hersel U, Dahmen C, Kessler H. 2003; RGD modified polymers: biomaterials for stimu-lated cell adhesion and beyond. Biomate-rials 24(24): 4385–4415.
ItoT, Itakura S, Todorov I et al. 2010; Mesenchy-mal stem cell and islet co-transplantation promotes graft revascularization and func-tion. Transplantation 89(12): 1438–1445. Karaoz E, Aksoy A, Ayhan S et al. 2009;
Char-acterization of mesenchymal stem cells
from rat bone marrow: ultrastructural properties, differentiation potential and immunophenotypic markers. Histochem Cell Biol 132(5): 533–546.
Karaoz E, Genc ZS, Demircan PC et al. 2010; Protection of rat pancreatic islet function and viability by coculture with rat bone marrow-derived mesenchymal stem cells. Cell Death Dis 1: e36.
Kerby A, Jones ES, Jones PM et al. 2013; Co-transplantation of islets with mesenchymal stem cells in microcapsules demonstrates graft outcome can be improved in an isolated-graft model of islet transplanta-tion in mice. Cytotherapy 15(2): 192–200. Kizilel S. 2010; Mathematical model for mi-croencapsulation of pancreatic islets within a biofunctional PEG hydrogel. Macromol Theory Simulat 19(8–9): 514–531. Kizilel R, Kizilel S. 2012; Application of the
numerical fractionation approach to the design of biofunctional PEG hydrogel membranes. Macromol React Eng 6(4): 160–179.
Kizilel S, Garfinkel M, Opara E. 2005; The bioartificial pancreas: progress and chal-lenges. Diabetes Technol Ther 7(6): 968–985. Kizilel S, Scavone A, Liu X et al. 2010; Encap-sulation of pancreatic islets within nano-thin functional polyethylene glycol coat-ings for enhanced insulin secretion. Tissue Eng A 16(7): 2217–2228.
Lee ST, Yun JI, Jo YS et al. 2010; Engineering integrin signaling for promoting embryonic
stem cell self-renewal in a precisely defined niche. Biomaterials 31(6): 1219–1226. Lee ST, Yun JI, van der Vlies AJ et al. 2012;
Long-term maintenance of mouse embryonic stem cell pluripotency by manipulating integrin sig-naling within 3D scaffolds without active Stat3. Biomaterials 33(35): 8934–8942. Li Y, Zheng X, Tang L et al. 2011; GLP-1
ana-logs containing disulfide bond exhibited prolonged half-life in vivo than GLP-1. Pep-tides 32(6): 1303–1312.
Liao SW, Rawson J, Omori K et al. 2013; Maintaining functional islets through en-capsulation in an injectable saccharide– peptide hydrogel. Biomaterials 34(16): 3984–3991.
Lin CC, Anseth KS. 2009; Glucagon-like peptide-1 functionalized PEG hydrogels promote survival and function of encapsu-lated pancreaticβ-cells. Biomacromolecules 10(9): 2460–2467.
Lin CC, Anseth KS. 2011; Cell–cell communi-cation mimicry with poly(ethylene glycol) hydrogels for enhancingβ-cell function. Proc Natl Acad Sci U S A 108(16): 6380–6385. Lin CC, Boyer PD, Aimetti AA et al. 2010;
Regulating MCP-1 diffusion in affinity hydrogels for enhancing immuno-isolation. J Control Release 142(3): 384–391. Lu Y, Jin X, Chen Y et al. 2010; Mesenchymal
stem cells protect islets from hypoxia/reoxygenation-induced injury. Cell Biochem Funct 28(8): 637–643. Mapelli C, Natarajan SI, Meyer JP et al. 2009;
Eleven amino acid glucagon-like peptide-1 receptor agonists with antidiabetic activity. J Med Chem 52(23): 7788–7799. Marek N, Krzystyniak A, Ergenc I et al. 2011;
Coating human pancreatic islets with CD4+CD25highCD127 regulatory T cells as a novel approach for the local immunoprotection. Ann Surg 254(3): 512–518; discussion, 518–519.
Matsumoto S. 2010; Islet cell transplantation for type 1 diabetes. J Diabetes 2(1): 16–22.
Mueller KR, Balamurugan AN, Cline GW et al. 2013; Differences in glucose-stimulated insulin secretion in vitro of is-lets from human, nonhuman primate, and porcine origin. Xenotransplantation 20(2): 75–81.
Muraglia A, Cancedda R, Quarto R. 2000; Clonal mesenchymal progenitors from hu-man bone marrow differentiate in vitro ac-cording to a hierarchical model. J Cell Sci 113(7): 1161–1166.
Nikolova G, Jabs N, Konstantinova I et al. 2006; The vascular basement mem-brane: a niche for insulin gene expres-sion and β cell proliferation. Dev Cell 10(3): 397–405.
Nuttelman CR, Tripodi MC, Anseth KS. 2005; Synthetic hydrogel niches that promote hMSC viability. Matrix Biol 24(3): 208–218. Omer A, Duvivier-Kali VF, Aschenbach W et al. 2004; Exercise induces hypoglycemia in rats with islet transplantation. Diabetes 53(2): 360–365.
O’Sullivan ES, Vegas A, Anderson DG et al. 2011; Islets transplanted in immuno-isolation devices: a review of the progress and the challenges that remain. Endocr Rev 32(6): 827–844.
Park KS, Kim YS, Kim JH et al. 2010; Trophic molecules derived from human mesenchy-mal stem cells enhance survival, function, and angiogenesis of isolated islets after transplantation. Transplantation 89(5): 509–517.
Sanz C, Vazquez P, Blazquez C et al. 2010; Signaling and biological effects of glucagon-like peptide 1 on the differentia-tion of mesenchymal stem cells from hu-man bone marrow. Am J Physiol Endocrinol Metab 298(3): E634–E643. Sims E, Evans-Molina C. 2012; Stem cells as
a tool to improve outcomes of islet trans-plantation. J Transpl 2012: 736491. Sreejalekshmi KG, Nair PD. 2011;
Biomimeticity in tissue engineering
scaffolds through synthetic peptide modi fi-cations– altering chemistry for enhanced biological response. J Biomed Mater Res A 96(2): 477–491.
Teramura Y, Iwata H. 2010; Bioartificial pan-creas microencapsulation and conformal coating of islet of Langerhans. Adv Drug Deliv Rev 62(7–8): 827–840.
Teramura Y, Oommen OP, Olerud J et al. 2013; Microencapsulation of cells, includ-ing islets, within stable ultra-thin mem-branes of maleimide-conjugated PEG– lipid with multifunctional crosslinkers. Biomaterials 34(11): 2683–2693. Vahl TP, Paty BW, Fuller BD et al. 2003;
Effects of GLP-1-(7–36)NH2, GLP-1-(7– 37), and GLP-1- (9–36)NH2 on intrave-nous glucose tolerance and glucose-induced insulin secretion in healthy humans. J Clin Endocrinol Metab 88(4): 1772–1779.
Weber LM, Anseth KS. 2008; Hydrogel en-capsulation environments functionalized with extracellular matrix interactions in-crease islet insulin secretion. Matrix Biol 27(8): 667–673.
Weber LM, Hayda KN, Haskins K et al. 2007; The effects of cell–matrix interactions on encapsulated β-cell function within hydrogels functionalized with matrix-derived adhesive peptides. Biomaterials 28(19): 3004–3011.
Weber LM, Hayda KN, Anseth KS. 2008; Cell–matrix interactions improve β-cell survival and insulin secretion in three-dimensional culture. Tissue Eng A 14(12): 1959–1968.
Yeung TY, Seeberger KL, Kin T et al. 2012; Human mesenchymal stem cells protect human islets from pro-inflammatory cyto-kines. PLoS One 7(5): e38189.
Zhu W, Xu W, Jiang R et al. 2006; Mesenchy-mal stem cells derived from bone marrow favor tumor cell growth in vivo. Exp Mol Pathol 80(3): 267–274.
![Figure 3. Optical microcope images of islets (20 islets/hydrogel) or islets + MSCs (20 islets + 20 000 MSCs/hydrogel) encapsulated within ligand functional PEG hydrogel scaffold ([IKVAV] = 2.5 m M and [GLP-1] = 20 μ M )](https://thumb-eu.123doks.com/thumbv2/9libnet/3973454.52539/6.892.152.744.97.366/optical-microcope-hydrogel-hydrogel-encapsulated-functional-hydrogel-scaffold.webp)