Amino acid, mineral, condensed tannin, and other chemical contents of olive leaves (Olea europaea L.) processed via solid-state fermentation using selected Aspergillus niger strains
Tam metin
Şekil
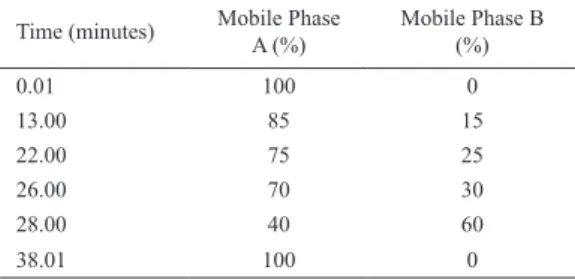
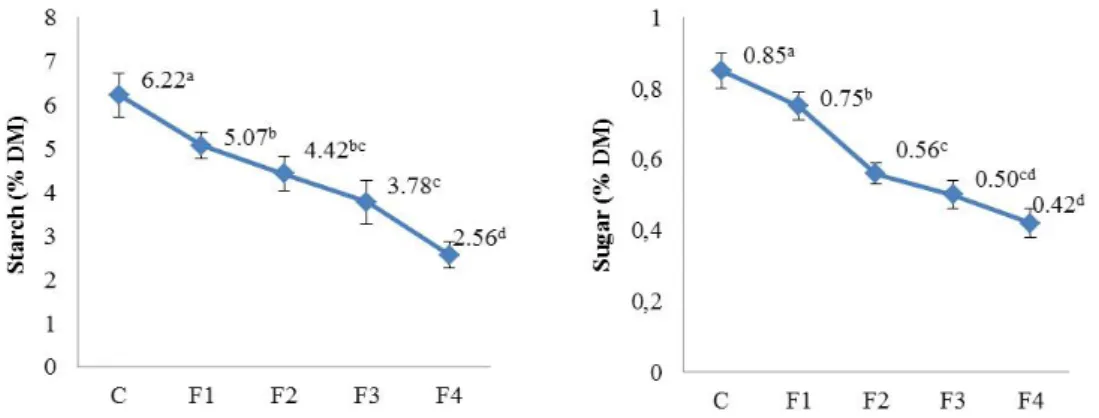
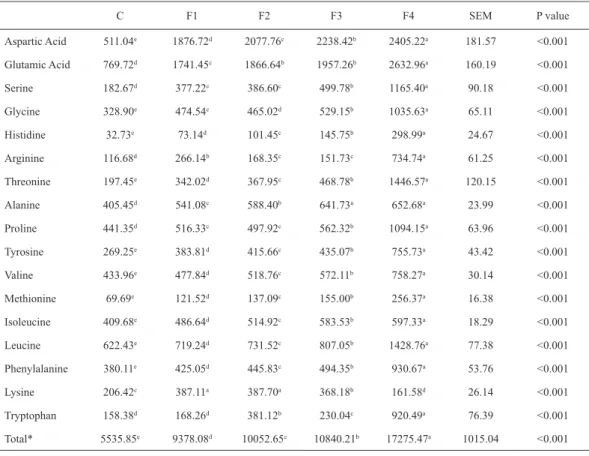
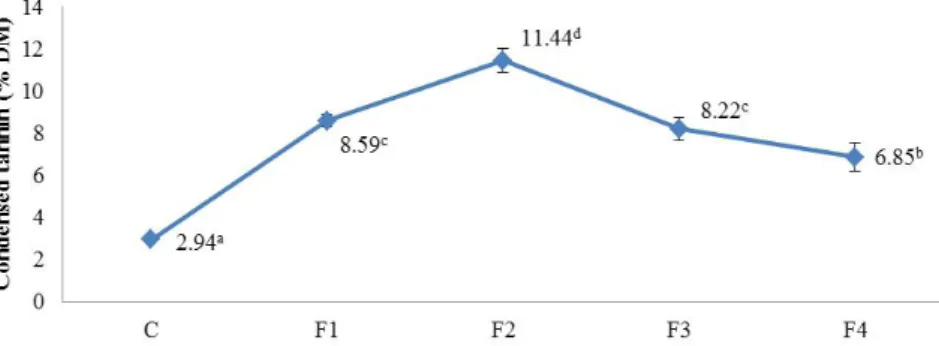
Benzer Belgeler
Daha sonraları CFC’lerle birlikte bu soğutkanlara alternatif olarak üretilen HCFC ve HFC türü halokarbon soğut- kanların da küresel ısınmaya neden oldukları ortaya
Tablo1’de yer alan araştırmaların sonuçlarına göre, tüketicilerin izinli mobil reklam mesajlarını kabul etme, izinsiz olarak gönderilen mobil reklamları ise kabul
20 ve 40’ı çıkarılmıştır. Aralamadan beş yıl sonra yapılan değerlendirmede, toplam meşcere hacmi aralanmış meşcerelerde kontrole göre düşerken, aralama
Although misophonia has not yet been defined as an effective treatment modality, cognitive beha- vioral psychotherapy, which is effectively used in the treatment of many
Olgu Sunumu: Otuz yaşında hemofili-C tanısı olan erkek hasta karın ağrısı, bulantı ve kusma şikâyetleri nedeniyle tarafımıza başvurdu.. Yapılan tetkiklerde ileal
Bu noktada aslında yazara göre, Hâcer’in bir “muhacir” olması ve Kâbe’nin varoluşunun onun tecrübesi üzerine inşa edilişi unutulmuştur.. Tüm bu okuma, yazarın kitap
Ancak, Heidegger’in cümlele- rin tarihsel olarak derse ait olduğu ve gelecekte de o yargıların bu metinde olması gerektiği vurgusu dikkate alındığında, Heidegger’in
Hukuk felsefesi, etik, siyaset felsefesi ve özgürlük sorununun tartışıldığı beşinci bölümde Torun, ağırlıklı olarak özgürlük problemi ve farklı etik teorileri üzerinde
