Full Terms & Conditions of access and use can be found at
https://www.tandfonline.com/action/journalInformation?journalCode=tfac20
Food Additives & Contaminants: Part A
ISSN: 1944-0049 (Print) 1944-0057 (Online) Journal homepage: https://www.tandfonline.com/loi/tfac20
Mycotoxin production capability of Penicillium
roqueforti
in strains isolated from mould-ripened
traditional Turkish civil cheese
Songul Cakmakci, Mustafa Gurses, A. Adnan Hayaloglu, Bulent Cetin, Pinar
Sekerci & Elif Dagdemir
To cite this article: Songul Cakmakci, Mustafa Gurses, A. Adnan Hayaloglu, Bulent Cetin, Pinar Sekerci & Elif Dagdemir (2015) Mycotoxin production capability of Penicillium�roqueforti in strains isolated from mould-ripened traditional Turkish civil cheese, Food Additives & Contaminants: Part A, 32:2, 245-249, DOI: 10.1080/19440049.2014.997808
To link to this article: https://doi.org/10.1080/19440049.2014.997808
Published online: 12 Jan 2015.
Submit your article to this journal
Article views: 350
View related articles
View Crossmark data
Mycotoxin production capability of
Penicillium roqueforti in strains isolated from mould-ripened
traditional Turkish civil cheese
Songul Cakmakcia*, Mustafa Gursesa, A. Adnan Hayaloglub, Bulent Cetina, Pinar Sekercia,cand Elif Dagdemira
a
Department of Food Engineering, Atatürk University, Erzurum, Turkey;bDepartment of Food Engineering,İnönü University, Malatya, Turkey;cDepartment of Food Engineering, Ardahan University, Ardahan, Turkey
(Received 7 September 2014; accepted 8 December 2014)
Mould-ripened civil is a traditional cheese produced mainly in eastern Turkey. The cheese is produced with a mixture of civil and whey curd cheeses (lor). This mixture is pressed into goat skins or plastic bags and is ripened for more than three months. Naturally occurring moulds grow on the surface and inside of the cheese during ripening. In this research, 140 Penicillium roqueforti strains were isolated from 41 samples of mould-ripened civil cheese collected from Erzurum and around towns in eastern Turkey. All strains were capable of mycotoxin production and were analysed using an HPLC method. It was established that all the strains (albeit at very low levels) produced roquefortine C, penicillic acid, mycophenolic acid and patulin. The amounts of toxins were in the ranges 0.4–47.0, 0.2–43.6, 0.1–23.1 and 0.1–2.3 mg kg−1, respectively. Patulin levels of the samples were lower than the others. The lowest level and highest total mycotoxin levels were determined as 1.2 and 70.1 mg kg−1respectively. The results of this preliminary study may help in the choice of secondary cultures for mould-ripened civil cheese and other mould-ripened cheeses.
Keywords: mould-ripened civil cheese; Penicillium roqueforti; mycotoxin; roquefortine C; Penicillic acid
Introduction
Civil cheese is a traditional cheese produced in eastern Turkey by using skimmed milk and the addition of whey in its manufacture. The cheese is produced by both rennet and acid coagulation and also a combination of heat treat-ments after coagulation of milk (Cakmakci 2011; Cakmakci et al. 2014). It is characterised by a fibrous structure which is formed by continuous stirring of milk during the coagulation period; and it has a low fat content and a plastic texture because of kneading during manu-facture (Figure 1, left). Civil cheese has a protected geo-graphical indication status and is produced in Erzurum in eastern Turkey and surrounding towns specified in docu-ments by Turkish Patent Institute (TPI) (2009). Because of its low-fat and high-protein content, its popularity has increased recently in terms of its nutrition composition (Cambaztepe et al. 2009). Another type of the cheese is manufactured by deliberate moulding of plain civil cheese and is called a mould-ripened civil cheese. In the produc-tion of this mould-ripened type of cheese, plain civil cheese is pressed into goat skins or plastic bags and ripened for more than three months (Cakmakci et al. 2012). It is dry-salted followed by ripening usually in hardened plastic barrels (Figure 1,middle). The cheese is spontaneously moulded during ripening and it has a dif-ferentflavour and appearance from the plain civil cheese. Mould-ripened cheese is preferred by people living around Erzurum and neighbouring cities (Figure 1, right).
However, the mould growth on the cheese surface may be a potential risk for public health, so research should be conducted to investigate this potential problem.
Mycotoxins are toxic secondary metabolites that are produced by many species of moulds (Bullerman 1981; Kokkonen et al.2005a). Penicillium species are ubiqui-tous, being found in air, soil, food and feed products (Houbraken et al. 2010; Visagie et al. 2014). It was reported that various mycotoxins have been produced on culture media by some of the fungal starters used in the manufacture of cheese (Kokkonen et al.2005a). The main beneficial property of these species is its role in the production of blue cheeses such as Roquefort, gorgon-zola and Danish blue (Gripon1993; Nichol2000; Geisen et al.2001; Tiwary et al.2009). Penicillium roqueforti is well adapted to cheese and contributes to the flavour development of blue-type cheeses through lipolytic and proteolytic enzymes (Geisen et al. 2001). It has been reported that P. roqueforti is a predominant mould spe-cies in many types of mould-ripened cheeses such as Roquefort, stilton, gorgonzola, tulum, cabrales, Danish blue and mould-ripened civil (Bullerman1981; O’Brien et al.2004; Cakmakci et al. 2012; Hymery et al. 2014). The species are known as potential mycotoxin producers when used in cheese manufacturing applications and under storage conditions, even though they only produce very low concentrations (Bullerman1981; Hymery et al.
2014).
*Corresponding author. Emails:[email protected];[email protected]
Vol. 32, No. 2, 245–249, http://dx.doi.org/10.1080/19440049.2014.997808
Various food-borne Penicillium species can be identified with traditional (morphological) and molecular (DNA-based) techniques. The technological properties of any given P. roqueforti strain are determined by their genetic make-up (Geisen et al.2001). In particular, Penicillium species can be identified based on examination of macroscopic, microscopic and morphological characteristics (e.g., colour, size, and shape of colonies and conidia) (Hasenekoğlu 1991; Samson et al. 2000; Cakmakci et al.2012).
Some strains of P. roqueforti are suitable for preparing a secondary starter for different blue cheese varieties. No studies are available on the identification of the indigenous fungi grown on these local blue cheeses. Since these cheeses are made by traditional procedures without inocu-lation with a pure culture (at difference of the industrial procedures), fungi other than P. roqueforti might contribute to their maturation. Indeed, P. roqueforti is closely related to some other Penicillium species that grow in foods (Boysen et al. 2000), e.g. P. paneum and P. carneum (Nielsen et al.2006). P. roqueforti and the related species produce several mycotoxins and other non-toxic secondary metabolites (Fernández-Bodega et al.2009). Among other metabolites, P. roqueforti produces PR toxin (Jelen et al. 2002), the alkaloid roquefortine C (a dimethylallyl-trypto-phan derivative) (Finoli et al.2001; Tiwary et al.2009), and mycophenolic acid (MPA) (Schneweis et al.2000).
Mycotoxins are secondary metabolites of some fila-mentous fungi and are a structurally diverse group of mostly low molecular weight compounds (Peraica et al. 1999; Milicevic 2009). Aspergillus, Penicillium and Fusarium genera, which may develop on various foods and feeds under suitable temperature and humidity condi-tions, cause serious risks to human and animal health (Milicevic2009). Furthermore, it is known that mycotox-ins are resistant to industrial processing. It has been reported that different fungal growth conditions such as substrate composition and environmental factors (tem-perature, water activity, pH or oxygen concentration) may change mycotoxin production capacity (Kokkonen et al.2005b; Magan & Aldred2007). Selective extraction procedures will in some cases be necessary to obtain a full profile of secondary metabolites, and in ecology even in a small part of a colony such as an interaction zone. In both cases a simple and rapid screening method will be
necessary. Whereas a large number of HPLC methods have been described for specific determination of metabo-lites, only a few methods are suited for general screening of cultures (Smedsgaard 1997).
This paper reports the mycotoxin production capabil-ities of isolated P. roqueforti strains (Cakmakci et al. 2012) spontaneously growing on traditional mould-ripened civil cheese and the accompanying risk for human health. In this context, the mycotoxin-producing capability of P. roqueforti was established by determining the presence of some mycotoxins including roquefortine C, penicillic acid, MPA and patulin in mould-ripened civil cheeses. In addition, it was intended to provide guidance in the selection of non-toxic P. roqueforti strains in this cheese and their use as their starter culture.
Materials and methods Cheese samples
Forty-one mould-ripened civil cheese samples were col-lected from retail outlets in Erzurum province (Turkey) and kept cold (4 ± 1°C) in sterile plastic bags during transport to the Research Laboratory at the Food Engineering Department, Atatürk University in Erzurum, Turkey. Roquefortine C (Santa Cruz Biotechnology Co. Ltd, Shanghai, China), patulin, MPA and penicillic acid (Sigma-Aldrich, St. Louis, MO, USA) were used as standards.
Growth media and conditions
Potato dextrose agar (PDA) (Merck, Darmstadt, Germany) (Figure 2), Czapex Dox agar (CDA; Merck) and yeast extract sucrose (YES) agar (Merck) (Figure 3) were used for the isolation of fungi. PDA was acidified with 10% tartaric acid to pH 3.5. Cheese (10 g) was dissolved in a 90 ml of 0.1% peptone in 0.85% NaCl solution in a sterile polyethylene bag and homogenised with a stomacher (HG 400, Mayo International, Milan, Italy) for 2 min. Serial dilutions of each cheese sample were prepared and inoculated onto PDA and CDA. The plates were incubated at 25°C in darkness for 7 days, as described by Harrigan (1998). Isolated fungal colonies Figure 1. (colour online) (left) Fresh civil cheese, (middle) civil cheese about to be pressed into its packaging and (right) mould-ripened civil cheese.
(140 strains) were identified using traditional and mole-cular methods (Cakmakci et al. 2012). Macroscopic and microscopic characteristics of the strains were examined using methods described by Hasenekoğlu (1991), Samson et al. (2000), and Samson and Frisvad (2004). In this study, toxins produced by P. roqueforti were determined using an HPLC method.
Secondary metabolite extraction from cultures and conditions for HPLC analysis
The extraction of toxin produced in vitro by P. roqueforti cultures from mould-ripened civil cheeses was prepared by the method described by O’Brien et al. (2006). Culture extracts were prepared by cutting three plugs of 6 mm diameter from several colonies using a cork drill as fol-lows (Figure 3). The plugs were transferred to a 4 ml disposable autosampler screw-cap vial and 500 µl of the
solvent mixture methanol-dichloromethane/ethyl acetate (1:2:3, v/v/v) containing 1% (v/v) formic acid were added. The plugs were extracted overnight (O’Brien et al. 2006). Extracts were prepared using the method described by Smedsgaard (1997). Each extract wasfiltered in a vial of 2 ml through a 0.45 µm PTFEfilter after 1–2 h and later analysed using HPLC with diode array detection (DAD) (Houbraken et al. 2010). A Shimadzu LC-20AD Prominence HPLC system (Shimadzu Corp., Kyoto, Japan) was used; the system consisted of a SPDM20A diode array detector, a SIL-20A HT autosampler, a CTO-20A column oven and a DGU-20A5 degasser. A Phenomenex Luna C18(2) column 100 × 4.6 mm × 3 um (Phenomenex, Torrance, CA, USA) was used for separa-tion. A gradient solvent system – solvent A and B were water and acetonitrile in a %0.05 (v/v) trifloroacetic acid, respectively– was used as the mobile phase at a flow rate of 0.5 ml min–1. Initially, the elution was started at 5% B and then with a gradient ranging from 5% to 100% over 30 min, maintained at 100% B for 5 min and followed by equilibration for 5% B for the next injection. The injection volume was 3 µl and oven temperature was 30°C. The elution was monitored at 274 nm for patulin and at 210 nm for penicillic acid, roquefortine C and MPA. The concentrations of mycotoxins in the extracts were calcu-lated by the external standard method. Mycotoxins were identified by the matching the retention time of the exter-nal standard. Also, UV spectra of the peaks of the myco-toxins were also matched with the relevant data from the literature.
Results and discussion
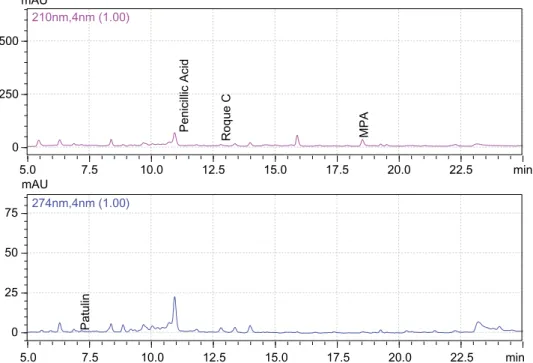
One hundred and forty strains of P. roqueforti isolated from 41 samples of mould-ripened civil cheeses were analysed for mycotoxin-forming capacity. The minimum, maximum and average values (mg kg−1) of secondary metabolites (roquefortine C, penicillic acid, MPA and patulin) formed are givenTable 1. Also, a representative HPLC-DAD chro-matogram is shown in Figure 4. The analytical results indicate that the all strains of P. roqueforti tested in this study were able to produce secondary metabolites at differ-ent levels in vitro. The lowest level and highest total myco-toxin levels were determined as 1.2 and 70.1 mg kg−1 respectively.
Figure 2. (colour online) Penicillium roqueforti from potato dex-trose agar (PDA).
Figure 3. (colour online) Culture extraction from yeast extract sucrose (YES) agar.
Table 1. Mycotoxin-producing potentials of Penicillium roque-forti strains isolated from mould-ripened civil cheeses (mg kg−1) (n = 140 P. roqueforti strains).
Mycotoxins Minimum Maximum Average
Patulin 0.1 2.3 0.6
Mycophenolic acid 0.1 23.1 3.6
Roquefortine C 0.2 47.0 3.6
Roquefortine C, penicillic acid and MPA showed var-iation in levels depending on the strain. As seen from
Table 1, roquefortine C was always found in the tested
isolates at levels ranging from 0.2 to 47 mg kg−1, aver-aging 3.6 mg kg−1. The consumption of blue cheese with relatively low levels of toxins or low toxicity of roquefor-tine may be safe for consumers (Finoli et al. 2001). Frisvad et al. (2004) reported that the most widespread mycotoxin was roquefortine C, which was produced by 25 of 58 Penicillium species. Moreover, Vinokurova et al. (2001) reported that roquefortine C and its precursor roquefortine D were produced by virtually all studied strains within the P. roqueforti species. Finoli et al. (2001) isolated the strains from the foreign cheeses that belonged to P. roqueforti and these strains were able to produce the roquefortine at between 0.18 and 8.44 mg l–1 in yeast extract sucrose medium. It was determined by Vallone et al. (2014) that only two strains out of nine produced roquefortine C in vitro. Hymery et al. (2014) reported that to date there have been no toxicity studies performed for roquefortine C on experimental animals that meet good laboratory practice standards. Rasmussen et al. (2011) indicated that eight fungal species significantly affected Caco-2 cell viability in vitro and that the 50% inhibition concentration (IC50) was 48μg ml–1for roque-fortine C. According to a review by Hymery et al. (2014) on mycotoxin forming in cheese, no recent data are avail-able for the estimation of human exposure.
Penicillic acid levels ranged from 0.2 to 43.6 mg kg−1, with an average of 5.2 mg kg−1 (Table 1). Erdogan
et al. (2003) also found using TLC that P. roqueforti strains produced high levels of penicillic acid in tulum cheeses. MPA acid and patulin amounts were 0.1– 23.1 mg kg−1, averaging 3.6 mg kg−1, and 0.1– 2.3 mg kg−1, averaging 0.6 mg kg−1, respectively
(Table 1). Vallone et al. (2014) reported that MPA was
produced by four of nine strains in vitro. Patulin was the secondary metabolite produced at the lowest con-centrations by P. roqueforti isolates (Table 1). These results tend to indicate that P. roqueforti is not a patulin producer. This is confirmed by Nielsen et al. (2006) who reported that patulin was the only metabolite pro-duced by P. paneum and P. carneum but that it was not produced by P. roqueforti.
In conclusion, the dominantfilamentous fungal species detected in mould-ripened civil cheese was P. roqueforti, and all these isolated strains were capable of producing second-ary metabolites (roquefortine C, penicillic acid, MPA and patulin) at different levels. For strict control of food quality, it is therefore necessary across the world to avoid such toxico-logical risk. This study has identified the secondary metabo-lites produced in vitro by P. roqueforti isolated from mould-ripened civil cheese. The results of this preliminary study may help determine the secondary cultures for mould-ripened civil cheese and other mould-mould-ripened cheeses. However, further research should be undertaken to look for the presence of additional mycotoxins other than these four mycotoxins studied here in cheese media. The toxin-forming capacity of moulds may change due to conditions in the environment and during cheese ripening.
5.0 7.5 10.0 12.5 15.0 17.5 20.0 22.5 min 0 250 500 mAU 210nm,4nm (1.00) Penicillic Acid Roque C MPA 5.0 7.5 10.0 12.5 15.0 17.5 20.0 22.5 min 0 25 50 75 mAU 274nm,4nm (1.00) Patulin
Figure 4. (colour online) HPLC-DAD chromatogram eluted at 210 nm for penicillic acid, roquefortine C and mycophenolic acid (top) and at 274 nm for patulin (bottom).
Funding
The authors thank the Scientific and Technological Research Council of Turkey (TÜBİTAK, Turkey) for supporting this study [TOVAG-Project numbers 108 O 509 and 111 O 199].
References
Boysen ME, Jacobsson K-G, Schnurer J. 2000. Molecular iden-tification of species from the Penicillium roqueforti group associated with spoiled animal feed. Appl Environ Microbiol. 66:1523–1526.
Bullerman LB. 1981. Public health significance of molds and mycotoxins in fermented dairy products. J Dairy Sci. 64:2439–2452.
Cakmakci S. 2011. Türkiye peynirleri. In: Hayaloglu AA, Ozer B, editors. Peynir Biliminin Temelleri. Izmir: Sidas; p. 585–614.
Cakmakci S, Cetin B, Gurses M, Dagdemir E, Hayaloglu AA. 2012. Morphological, molecular, and mycotoxigenic identi-fication of dominant filamentous fungi from moldy civil cheese. J Food Prot. 75:2045–2049.
Cakmakci S, Hayaloglu AA, Dagdemir E, Cetin B, Gurses M, Tahmas-Kahyaoglu D. 2014. Effect of Penicillium roqueforti and whey cheese on gross composition, microbiology and proteolysis of mould-ripened civil cheese during ripening. Int J Dairy Technol. 67:594–603.
Cambaztepe F, Cakmakci S, Dagdemir E. 2009. Effect of some technological parameters on microbiological, chemical and sensory qualities of civil cheese during ripening. Int J Dairy Technol. 62:541–548.
Erdogan A, Gurses M, Sert S. 2003. Isolation of moulds capable of producing mycotoxins from blue mouldy Tulum cheeses produced in Turkey. Int J Food Microbiol. 85:83–85. Fernández-Bodega MA, Mauriz E, Gómez A, Martín JF. 2009.
Proteolytic activity, mycotoxins and andrastin A in Penicillium roqueforti strains isolated from Cabrales, Valdeón and Bejes–Tresviso local varieties of blue-veined cheeses. Int J Food Microbiol. 136:18–25.
Finoli C, Vecchio A, Galli A, Dragoni I. 2001. Roquefortine C occurrence in blue cheese. J Food Prot. 64:246–251. Frisvad JC, Smedsgaard J, Larsen TO, Samson RA, Robert A.
2004. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud Mycol. 49:201–241.
Geisen R, Cantor MD, Hansen TK, Holzapfel WH, Jakobsen M. 2001. Characterization of Penicillium roqueforti strains used as cheese starter cultures by RAPD typing. Int J Food Microbiol. 65:183–191.
Gripon JC. 1993. Major cheese groups: mould-ripened cheeses. In: Fox PF, editor. Cheese: chemistry, physics and microbiology. Vol. 2. London: Chapman & Hall; p. 111–136.
Harrigan WF. 1998. Laboratory methods in food microbiology. San Diego, CA: Academic Press.
Hasenekoğlu İ. 1991. Toprak mikrofungusları (6 Cilt). Yay. No: 689. Erzurum: Atatürk University.
Houbraken J, Frisvad JC, Samson RA. 2010. Sex in Penicillium series Roqueforti. IMA Fungus. 1:171–180.
Hymery N, Vasseur V, Coton M, Mounier J, Jany J-L, Barbier G, Coton E. 2014. Filamentous fungi and mycotoxins in cheese: a review. Compr Rev Food Sci Food Saf. 13:437–456. Jelen HH, Mildner S, Czaczyk K. 2002. Influence of octanoic
acid addition to medium on some volatile compounds and
PR-toxin biosynthesis by Penicillium roqueforti. Lett Appl Microbiol. 35:37–41.
Kokkonen M, Jestoi M, Rizzo A. 2005a. Determination of selected mycotoxins in mould cheeses with liquid chromato-graphy coupled to tandem with mass spectrometry. Food Addit Contam. 22:449–456.
Kokkonen M, Jestoi M, Rizzo A. 2005b. The effect of substrate on mycotoxin production of selected Penicillium strains. Int J Food Microbiol. 99:207–214.
Magan N, Aldred D. 2007. Why do fungi produce mycotoxins? In: Dijkserhuis J, Samson RA, editors. Food mycology: a multifaceted approach to fungi and food. New York: CRC Press/Taylor and Francis Group; p. 121–133.
Milicevic D. 2009. Mycotoxins in the food chain– old problems and new solutions. Tehnologija Mesa. 50:99–111.
Nichol AW. 2000. Cheese/mould-ripened varieties. In: Robinson RK, editor. Encyclopedia of food microbiology. San Diego (CA): Academic Press; p. 387–393.
Nielsen KF, Sumarah MW, Frisvad JC, Miller JD. 2006. Production of metabolites from the Penicillium roqueforti complex. J Agric Food Chem. 54:3756–3763.
O’Brien M, Nielsen KF, O’Kiely P, Forristal PD, Fuller HT, Frisvad JC. 2006. Mycotoxins and other secondary metabo-lites produced in vitro by Penicillium paneum Frisvad and Penicillium roqueforti Thom isolated from baled grass silage in Ireland. J Agric Food Chem. 54:9268–9276.
O’Brien NM, O’Connor TP, O’Callaghan J, Dobson ADW. 2004. Toxins in cheese biogenic amines and mycotoxins in cheese: chemistry, physics and microbiology. In: Fox PF, Guinee T, Cogan T, McSweeney P, editors. General aspects. 3rd ed. Vol. 1. Chapter 19, Amsterdam: Elsevier Applied Science. Peraica M, Radić B, Lucić A, Pavlović M. 1999. Toxic effects of
mycotoxins in humans. Bull World Health Organ. 77: 754–766.
Rasmussen RR, Rasmussen PH, Larsen TO, Bladt TT, Binderup ML. 2011. In vitro cytotoxicity of fungi spoiling maize silage. Food Chem Toxicol. 49:31–44.
Samson RA, Frisvad JC. 2004. Penicillium subgenus Penicillium: new taxonomic schemes, mycotoxins and other extrolites. Stud Mycol. 49:1–251.
Samson RA, Hoekstra ES, Frisvad JC, Filtenborg O. 2000. Introduction to food- and airborne fungi. 6th ed. Utrecht, The Netherlands: Centraalbureau voor Schimmelcultures. Schneweis I, Meyer K, Hormansdorfer S, Bauer J. 2000.
Mycophenolic acid in silage. Appl Environ Microbiol. 66:3639–3641.
Smedsgaard J. 1997. Micro-scale extraction procedure for stan-dardized screening of fungal metabolite production in cul-tures. J Chromatogr. 760:264–270.
Tiwary AK, Puschner B, Poppenga RH. 2009. Using roquefor-tine C as a biomarker for penitrem A intoxication. J Vet Diagn Investig. 21:237–239.
TPI. 2009. Civil Peynir Coğrafi İşaret Tescil Belgesi. Ankara: Turkish Patent Institute.
Vallone L, Giardini A, Soncini G. 2014. Secondary metabolites from Penicillium roqueforti, a starter for the production of Gorgonzola cheese. Italian J Food Saf. 3:173–177.
Vinokurova NG, Boichenko DM, Baskunov BP, Zelenkova NF, Vepritskaya IG, Arinbasarov MU, Reshetilova TA. 2001. Minor alkaloids of the fungus Penicillium roqueforti Thom 1906. Appl Biochemistry Microbiol. 37:184–187.
Visagie CM, Houbraken J, Frisvad JC, Hong S-B, Klaassen CHW, Perrone G, Seifert KA, Varga J, Yaguchi T, Samson RA. 2014. Identification and nomenclature of the genus Penicillium. Stud Mycol. 78:343–371.