1993
BAŞKENT ÜNİVERSİTESİ
TIP FAKÜLTESİ
İç Hastalıkları Anabilim Dalı
Endokrinoloji ve Metabolizma Bilim Dalı
HİPERPROLAKTİNEMİK OLGULARDA
PROLAKTİN-MAKROPROLAKTİN VE ADP İLE İNDÜKLENMİŞ TROMBOSİT
AKTİVASYONU İLİŞKİSİ
YANDAL UZMANLIK TEZİ
Uz. Dr. İnan ANAFOROĞLU
1993
BAŞKENT ÜNİVERSİTESİ
TIP FAKÜLTESİ
İç Hastalıkları Anabilim Dalı
Endokrinoloji ve Metabolizma Bilim Dalı
HİPERPROLAKTİNEMİK OLGULARDA
PROLAKTİN-MAKROPROLAKTİN VE ADP İLE İNDÜKLENMİŞ TROMBOSİT
AKTİVASYONU İLİŞKİSİ
YANDAL UZMANLIK TEZİ
Uz. Dr. İnan ANAFOROĞLU
Tez Danışmanı: Doç. Dr. Nilgün GÜVENER DEMİRAĞ
Proje No: KA 05/85
TEŞEKKÜR
Endokrinoloji ve Metabolizma Hastalıkları eğitimimi en iyi şekilde tamamlamamı sağlamak için yapmış oldukları çok değerli katkılarından dolayı başta Sayın rektörümüz Prof. Dr. Mehmet Haberal olmak üzere, Dahili Tıp Bilimleri Başkanı Sayın Prof. Dr. Haldun Müderrisoğlu’na, İç Hastalıkları Anabilim Dalı Başkanı Sayın Prof. Dr. Nurhan Özdemir’e, Endokrinoloji ve Metabolizma Bilim Dalı Başkanı ve aynı zamanda, tezimin her aşamasında büyük emeği olan tez danışmanım, Sayın Doç. Dr. Nilgün Güvener Demirağ’a ve Doç. Dr. Neslihan Başçıl Tütütncü’ye şükranlarımı sunarım.
Eğitimimin tamamlanmasında büyük katkıları olan Endokrinoloji ve Metabolizma Bilim Dalı öğretim üyeleri, Yrd. Doç. Dr. M. Eda Ertörer, Yrd. Doç. Dr. Alptekin Gürsoy’a, Merkez Müdürümüz Sayın Yrd. Doç. Dr. Turgut Noyan’a, asistan arkadaşlarıma, sevgi ve desteklerini hep yanımda hissettiğim aileme çok teşekkür ederim.
ÖZET
Trombositlerin ADP ile stimülasyonunun, serum PRL düzeyleri ile pozitif korelasyon gösterdiği ve hiperprolaktineminin, trombosit agregasyonu için bir risk faktörü olabileceği son zamanlarda bildirilmiştir. Prolaktin yüksekliği olan hastalarda yapılan çalışmalar, hiperprolaktineminin, trombosit aktivasyonu için uygun bir ortam oluşturduğu bilinen ateroskleroz ve insülin direnci ile de bağlantılı olduğunu düşündürmektedir. Bu çalışmada, hiperprolaktinemi ile trombosit aktivasyonuna bağlı P-selektin ekspresyonu arasındaki ilişki, hastaların metabolik durumları da göz önünde bulundurularak irdelendi.
Trombosit fonksiyonlarını etkilediği bilinen tüm faktörler dışlandıktan sonra, premenopozal dönemde bulunan ve hiç tedavi almamış olan hiperprolaktinemik kadın olgular (n=32), yaş ve vücut kitle indeksleri benzer normoprolaktinemik olgular (n=33) ile karşılaştırıldı; yaş; 30,6±8 karşı 29,8±7,7 yıl, vücut kitle indeksi; 26,8±5,4 karşı 24,8±5,2 kg/m2, prolaktin 1889,8±886 karşı 335,9±117,9 mU/lt.
Gruplar arasında HOMA-IR (Homeostasis Model Assessment of Insulin Resistance) formülü ile hesaplanan insülin duyarlılığı, bel çevresi, kan basıncı, açlık plazma glukozu, insülini ve lipidler bakımından anlamlı fark olmaması şartı arandı. Trombositlerin ADP bağımlı P selektin ekspresyonunu göstermek için flovsitometri yöntemi kullanıldı. Hiperprolaktinemik grupta serum prolaktin düzeyleri polietilen glikolle (PEG) muamele edilmeden ve edildikten sonra ölçüldü. PEG ile muameleden sonra %40’ın üzerinde çökme olması durumu, makroprolaktin pozitifliği olarak kabul edildi.
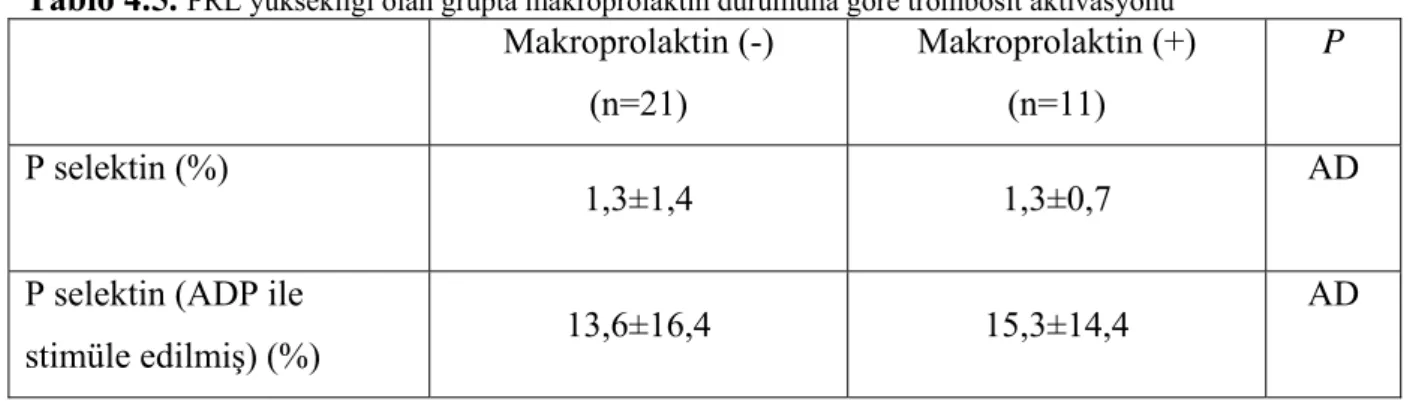
Trombositlerin ADP ile indüklenmiş P-selektin ekspresyonu hiperprolaktinemik grupta daha fazlaydı; [%14,2±15,5 karşı %6,7±5,2, (p=0.01) ]. Makroprolaktinemi sıklığı %29 bulundu. Prolaktin düzeyleri ve ADP ile indüklenmiş P selektin ekspresyonları arasında anlamlı korelasyon vardı (r=0,3, p<0,02). P selektin ekspresyonu makroprolaktin negatif (gerçek hiperprolaktinemi) (n=21) ve makroprolaktin pozitif (n=11) grupta benzerdi; %13,6±16,4 karşı %15,3±14,4 (p=0,7). P selektin ekspresyonunun, her iki alt-grupta kontrol grubundan yüksek olduğu izlendi; 513,6±16,4 (p=0,03) ve %15,3±14,4, (p=0,005), sırası ile.
Trombosit aktivasyonu, insülin direnci ile ilişkili olarak aterosklerotik hastalıkların patogenezinde rol alır. Bu çalışmada, insülin duyarlılığı belirteçleri bakımından tamamen benzer olan iki grupta, hiperprolaktineminin kendisinin trombosit aktivasyonu için bir risk faktörü olduğu ortaya konuldu. Aynı zamanda, makroprolaktineminin de, tıpkı gerçek hiperprolaktinemik hastalarda olduğu gibi, artmış trombosit aktivasyonu ile ilişkili olduğu gösterildi.
İNGİLİZCE ÖZET
Platelet activation is a recently recognized characteristic of prolactin, which functions through the potentiation of ADP-induced P selectin expression on platelets. Studies in hyperprolactinemic patients demonstrated atherosclerotic disorders related to insulin resistance; suitable milieus for platelet activation. We studied on hyperprolactinemic and normoprolactinemic patients to investigate the association between hyperprolactinemia and platelet activation related to P selectin expression,
After exclusion of any factor that might interfere with platelet functions, 32 naïve hyperprolactinemic and 33 age-body mass index-matched normoprolactinemic, non-smoking premenopausal women were included; age: 30.6±8 vs 29.8±7.7 years, body mass index: 26.8±5.4 vs 24.8±5.2 kg/m2, prolactin 1889.8±886 vs 335.9±117.9 mU/lt. Measurements regarding insulin sensitivity; which included, waist circumference, blood pressure, fasting plasma glucose, insulin and lipids were also matched. The flow-cytometry method was used to determine the ADP stimulated P selectin expression of the platelets. Serum prolactin was measured before and after polyethylene glycol precipitation (PEG) in hyperprolactinemic group. The diagnosis of macroprolactinemia was regarded as certain if the prolactin recovery in a serum was <40%.
The ADP stimulated P-selectin expression of the platelets was higher in hyperprolactinemic group; 14.2±15.5% vs 6.7±5.2%, (p=0.01). The frequency of macroprolactinemia was found to be 29%. There was a significant correlation between prolactin levels and ADP stimulated P selectin expression before PEG (r=0.3, p<0.02). The ADP stimulated P selectin expression rates were similar between macroprolactin negative (true hyperprolactinemia) (n=21) and macroprolactin positive (n=11) subgroups; 13.6±16.4% vs 15.3±14.4% (p=0.7). It kept being higher than the controls in both subgroups; 13.6±16.4%, (p=0.03) and 15.3±14.4%, (p=0.005), respectively.
Platelet activation is involved in the pathogenesis of atherosclerotic disorders related to insulin resistance. In this study, performed on completely matched group of cases regarding insulin sensitivity markers, hyperprolactinemia itself has been detected to bring an increased risk for platelet activation. It has also been clearly demonstrated that macroprolactinemia may cause platelet aggregation just as true hyperprolactinemia.
İÇİNDEKİLER
Sayfa No
TEŞEKKÜR ………..iii
ÖZET VE ANAHTAR SÖZCÜKLER……….iv
İNGİLİZCE ÖZET (ABSTRACT AND KEY WORDS)………v
İÇİNDEKİLER………..vi KISALTMALAR DİZİNİ………vii TABLOLAR DİZİNİ……….ix ŞEKİLLER DİZİNİ…..………..x 1. GİRİŞ VE AMAÇ………1 2. GENEL BİLGİLER……….2 2.1. Prolaktin yapısı………...2
2.2. Prolaktin sekresyonunun düzenlenmesi………3
2.3. Prolaktin reseptörü………4
2.3.1. Yapısı ve dağılımı………4
2.3.2. Prolaktin reseptörü aktivasyonu………5
2.4. Prolaktinin fonksiyonları………..8
2.5. Prolaktin ve trombosit ilişkisi………...14
2.5.1. Trombosit aktivasyonu, agregasyonu………14
2.6. Hiperprolaktinemi……….16
2.6.1. Hipofiz kaynaklı………...16
2.6.2. Hipofiz dışı nedenler……….……16
2.6.3. Makroprolaktinemi……….…….17
2.7. Metabolik sendrom, insülin direnci, trombosit aktivasyonu……….…….19
2.7.1. Metabolik sendrom ve insülin direnci………..……...19
2.7.2. Metabolik sendrom, insülin direnci, prolaktin………..……….18
2.7.3. Metabolik sendrom, insülin direnci, trombosit aktivasyonu….…………20
3. HASTALAR VE YÖNTEM…...………..….……….21 3.1. Çalışma grubu………..……….21 3.2. Yöntem………..……….21 3.3. İstatistiksel değerlendirme………...……….23 4. BULGULAR………..……….24 5. TARTIŞMA……….………...30 6. KAYNAKLAR………37
KISALTMALAR DİZİNİ
PRL: ProlaktinkDa: kiloDalton Ig: İmmünglobulin
TİDA: Tüberoinfindibuler hücreler VİP: Vazoaktif intestinal peptid cAMP: siklik adenozin monofosfat TRH: Tiroid hormonu salgılatıcı hormon GHRH: Büyüme hormonu salgılatıcı faktör GnRH: Gonadotropin salgılatıcı hormon GH: Büyüme hormonu
PL: Plasental laktojen
PRLR: Prolaktinin reseptörü EPO: Eritropoetin
IL: İnterlökin Jak 2: Janus kinaz 2
Stat: Signal transducer and activator of transcription MAPK: Mitojen aktive edici protein kinaz
IRS-1: İnsülin reseptör sustratı 1 PI: Fosfatidil inozitol
CIS: Cytokine-Inducible SH2-containing family; SH2 içeren, sitokin ile indüklenebilen protein ailesi
SOCS: Suppressors Of Cytokine Signaling; Sitokin Sinyalizasyonunun Baskılayıcıları PLC: Phospholipase C; Fosfolipaz C
ATP: Adenozintrifosfat
FFA: Free Faty Acid; serbest yağ asidi LPL: lipoprotein lipaz
FAS: Fatty Acid Synthase; yağ asidi sentaz
IGF-1: Insulin like Growth Factor-1; insülin benzeri madde 1 EGF: Epidermal Growth Factor; epidermal büyüme faktörü GpIIb/IIa: Glikoprotein IIb/IIa
ADP: Adenozindifosfat VTE: Venöz tromboemboliler
PKOS: Polikistik over sendromu JFK: Jel filtrasyon kromotografisi PEG: Polietilenglikol
IDF: Uluslararası Diyabet Federasyonu VKİ: Vücut kitle indeksi
PAI-1: Plazminojen aktive edici faktör
CMIA: Kemiluminesan mikropartikül immunoassay HOMA-IR: Homeostasis Model Assesment
AD: Anlamlı değil
PES: Parsiyel empty sella
OGTT: Oral glukoz tolerans testi CRP: C-reaktif protein
TNF a: Tümör nekroz faktörü alfa MRG: Manyetik rezonans görüntüleme FSH: Folikül stimüle edici hormon LH: Luteinize edici hormon CVO: Serebrovasküler olay AKS: Akut koroner sendrom
ŞEKİLLER DİZİNİ
Şekil 3.1 Trombositlerin flovsitometrede seçilmesi….………..21 Şekil 4.1. Kontrol grubunda P selektin ekspresyonu….……….25 Şekil 4.2. Gerçek hiperprolaktinemi grubunda P selektin ekspresyonu…….26 Şekil 4.3. Makroprolaktin grubunda P selektin ekspresyonu……….26
TABLOLAR DİZİNİ
Tablo 2.1. IDF metabolik sendrom tanı kriterleri………18 Tablo 4.1. Hastaların ve kontrol grubunun karakteristik özellikleri ve laboratuvar verileri…23 Tablo 4.2. PRL yüksekliği olan hastalarda başvuru nedenleri……….24 Tablo 4.3. PRL yüksekliği olan hastalarda MR bulguları………...24 Tablo 4.4. Hastalar ve kontrol grubunun trombosit aktivasyonu……….27 Tablo 4.5. PRL yüksekliği olan grupta makroprolaktin durumuna göre trombosit aktivasyonu….…27 Tablo 4.6. Makroprolaktin (-) hastalarla, kontrol grubunda trombosit aktivasyonu…………27 Tablo 4.7. Makroprolaktin (+) hastalarla, kontrol grubunda trombosit aktivasyonu………...28
1. GİRİŞ VE AMAÇ
PRL, vücutta yaygın olarak reseptörleri bulunan ve pek çok dokuda eksprese edilen bir hormondur. Eskiden sanıldığı gibi, sadece gebelik ve laktasyonda salgılanmadığı, vücutta pek çok durumda miktarı arttığı ve pek çok sistemi etkilediği ortaya konmuştur. Su ve elektrolit dengesi, büyüme ve gelişme, endokrinoloji ve metabolizma, beyin ve davranış, üreme, immünregülasyon ve koruma ile ilgili fonksiyonları olduğu bilinmektedir. Biyolojik olarak inaktif olan, farklı molekül ağırlığındaki PRL’in kanda yükselmesi, makroprolaktinemi olarak adlandırılmaktadır. Klinik önemi ve hiperprolaktinemi tespit edilen her hastada bakılıp bakılmayacağı, gittikçe, daha çok gündeme gelen konulardır. Son yıllarda ortaya atılan bir görüş de, PRL’in, vücuttaki pek çok doku yanında, trombosit fonksiyonlarını da etkilediği ve trombosit aktivasyonunu artırdığı yönündedir. Trombosit aktivasyonu, özellikle trombotik olaylarda ve önemli bir mortalite ve morbidite nedeni olan aterosklerotik olaylarda artmaktadır. Çalışmamızda, trombosit aktivasyonu ile hiperprolaktinemi arasındaki ilişkiyi incelerken, makroprolaktineminin klinik önemini de vurgulamaya amaçladık.
2. GENEL BİLGİLER
2.1. Prolaktin yapısıProlaktin (PRL), başlıca, pre-prolaktin olarak adenohipofizdeki asidofilik, laktotrof hücrelerden sekrete edilir. PRL’in esas fonksiyonu, gebe ve doğum yapmış kadınlarda memelerin gelişimi ve büyümesini ve laktasyonun devamının sağlanmasıdır (1). İnsanlarda, PRL’in hipofiz dışında da, vücutta pek çok dokudan salgılandığı gösterilmiştir. Hem dolaşımda bulunarak, hem de otokrin veya parakrin etkileri ile, PRL’in üreme, metabolik, ozmoregülatuvar ve immünregülatuvar gibi sistemlere etkili bir hormon olduğu bilinmektedir (2-5).
İnsan PRL geni 6. kromozom üzerinde bulunur (6). PRL, 199 aminoasitten oluşan, 3 tane disülfid bağı olan, 26 kiloDalton (kDa) ağırlığında bir pre-hormon olarak sentez edilir (7). Pre-hormonun proteolitik olarak ayrılması sonucunda, 23 kDa ağırlığındaki, matür, monomerik form açığa çıkar. Normal kişilerin ve hiperprolaktinemisi olan kişilerin serumunda, %85-95 oranında monomerik form hakimdir. Küçük (Little) PRL de denilen monomerik PRL, biyolojik olarak en aktif formdur (7, 4). Dolaşımda, monomerik formun yanında değişik moleküler ağırlığa sahip olan farklı formlar da bulunur. Farklı formların bulunmasının; amino asid zincirlerinin düzenlenmesi sırasında alternatif bağlanma bölgelerinin oluşması, proteolitik ayrılma ve diğer translasyonel düzenlemelerin farklılığından kaynaklanabileceği düşünülmüştür (4). PRL mRNA’sının alternatif bağlanması ile ortaya çıktığı düşünülen 137 amino asitlik bir PRL varyantı hipofizde gösterilmiştir (7-9). Alternatif bağlanma, farklı PRL varyantları için major bir kaynak olarak görülmemektedir (4). Proteolitik ayrılmada çeşitli enzimlerin rol aldığı düşünülmektedir. En çok çalışılan formlar, 14-16-22 kDa ağırlığındaki moleküllerdir. Yapılan çalışmalar, ölçüm sırasında ve ölçüm yöntemlerine bağlı olarak da ortaya çıkabileceklerini düşündürdüğünden, gerçek fizyolojik önemleri bilinmemektedir (7). Alternatif bağlanma ve proteolitik ayrılma yanında, PRL formlarının önemli kısmı, dimerizasyon, polimerizasyon, fosforilasyon, glikolizasyon, sülfasyon, deamidasyon işlemlerini içeren diğer translasyonel düzenlemelerden kaynaklanmaktadır (4, 10). PRL’nin dimerizasyon ve polimerizasyonu ya da immünglobulin (Ig) gibi bağlayıcı proteinlerle kovalen veya non-kovalen agregasyonu sonucu makroprolaktin adı verilen yüksek ağırlıklı moleküller oluşur (7, 11-13). Dimerizasyon ve polimerizasyon işlemleri dışındaki işlemlerle gerçekleşen formlar insanlarda gösterilememiştir (4). ‘Büyük (Big) PRL’, 50 kDa ağırlığındaki dimerik PRL de denilen form, PRL moleküllerinin, dimerik,
kovalen bağlanmasından oluşmaktadır. Serumdaki PRL’nin %10-15’ini oluşturur. ‘Büyük büyük (Big big) PRL’ ya da makroprolaktin, serumdaki PRL’nin %1’inden azını oluşturur. Makroprolaktin, yaklaşık 150 kDa ağırlığındadır (14).
2.1. PRL sekresyonunun düzenlenmesi
PRL sekresyonu, tüberoinfindibuler hücreler (TİDA) ve hipotalamik tüberohipofzyel dopaminerjik sistemden salgılanan dopaminin inhibitör kontrolü altındadır (15, 16). Dopaminin, PRL sekresyonunu tam olarak hangi mekanizmalarla yaptığı net olmamakla birlikte, dopamin, hipotalamik hipofizer portal sistemden geçerek, laktotroplar üzerindeki D2 reseptörlerine bağlanarak PRL sekresyonunu inhibe eder (16). Saniyeler içinde, dopamin, potasyum iletimini artırarak, voltaj duyarlı kalsiyum kanallarını inaktive eder, membran hiperpolarizasyonu oluşur, hücre içi serbest kalsiyum düzeyi azalır. Dakikalar ile saatler içinde, dopamin, adenilat siklaz aktivitesini, inozitol fosfat metabolizmasını, araşidonik asit salınımını ve PRL gen ekspresyonunu baskılar. Günler içinde ise, dopamin, hücre proliferasyonunu baskılar. Bir paradoks olarak, çok düşük düzeylerdeki dopaminin PRL salınımını artırdığı gözlenmiştir (2). PRL, TİDA nöronlarındaki, tirozin hidroksilaz aktivitesini artırarak, salınımı üzerindeki negatif kontrol mekanizmasına katkıda bulunur (16). Dopamin dışında, PRL sekresyonunun düzenlenmesine katkıda bulunan endotelin-1, transforme edici büyüme faktörü ve kalsitonin parakrin PRL inhibitörleri olarak görev yaparlar (17-19). PRL sekresyonunu artıran faktörler de mevcuttur. Bazal fibroblast kaynaklı büyüme faktörü ve epidermal büyüme faktörü PRL sentez ve sekresyonunu artırır. Vazoaktif intestinal peptid (VİP), PRL sentezini siklik adenozin monofosfat (cAMP) üzerinden artırırken, oksitosin ve hipofizer adenilat siklaz aktive edici protein de PRL sekresyonunu artırır (17, 20). Medial bazal hipotalamustaki immünreaktif hücrelerin yaklaşık %33’ünün östrojenle işaretlenmesi, bu nöronlarda östrojen reseptörlerinin olduğuna işaret etmektedir (21). Ön hipofiz laktotrof hücrelerinin gelişimini kontrol eden transkripsiyon düzenleyicileri de erişkin hayatta PRL sentezinin düzenlenmesinde rol alır. Bu faktörler içinde en göze çarpanı Pit-1 proteinidir. Östrojen ve cAMP gibi transkripsiyon faktörleri Pit-1 aktivitesini artırarak, PRL gen ekspresyonunu artırırlar (16). Östrojenin PRL gen transkripsiyonunu ve sekresyonunu uyarması, kadınların PRL düzeylerinin erkeklere göre ve düzenli adet gören kadınların menopoz dönemindeki kadınlara göre neden daha yüksek olduğunu da açıklamaktadır (21, 22). Tiroid hormonu salgılatıcı hormon (TRH), PRL sekresyonunu uyarmakla birlikte, PRL’nin düzenli salınımında önemli bir rol oynamamaktadır (23, 24). Hem hipofizde, hem de hipotalamusta sentezlenen galanin, PRL salgılatıcı bir faktör olarak
kabul edilmektedir (20). Aminobütirik asit, nörotensin, substans P, bombesin ve kolesistokininin PRL sekresyonu üzerindeki etkileri net değildir (17). PRL salgılatmada, serotoninin VİP’e katkıda bulunduğu düşünülmektedir. Noktürnal PRL sekresyonu, siproheptadinle artmaktadır, buradan yola çıkarak, serotoninin noktürnal sekresyonu düzenlemede görev aldığı düşünülmektedir. Serotonin ve opiyatlar, akut olarak PRL salınımını artırır (16, 25). Büyüme hormonu salgılatıcı faktör (GHRH), yüksek dozlarda, ılımlı olarak PRL sekresyonunu artırmaktadır. GHRH üreten tümörü olan hastalarda PRL düzeyleri yüksek bulunmuştur. Gonadotropin salgılatıcı hormonun (GnRH) da, özellikle kadınlarda, periovulatuvar dönemde, PRL sekresyonunu artırdığı gözlenmiştir (25). Arka hipofiz hormonları olan vazopressin ve oksitosinin PRL salınımı üzerindeki etkileri net değildir. Histaminin, hipotalamus üzerinden PRL salınımını etkileyebileceği düşünülür, histamin reseptör blokerleri PRL sekresyonunu artırmaktadır (26).
PRL’in, gün içindeki normal sekresyon paterni, her 2-3 saatte bir hormonun bezden salınımı şeklindedir. En fazla hormon salgılanması ise, uykunun hızlı göz hareketlerinin olduğu dönemde gerçekleşmektedir, bu nedenle en yüksek salınım geceleri gerçekleşmektedir (27). Emziren kadınlarda, emme eylemi, PRL için güçlü bir uyarıcı olmaktadır. Emosyonel stres ve seksüel orgazm da PRL’i artıran diğer faktörlerdir. PRL sekresyonunu inhibe eden ve uyaran, yukarıda bahsedilen faktörlerin, PRL salgılatan bu olaylarla olan ilgisi net bilinmemektedir (16).
Sağlıklı kadınlarda, normal plazma PRL konsantrasyonu 4-20 ng/ml’dir. Erkeklerde bu değerler daha düşüktür (16).
2.3. Prolaktin reseptörü 2.3.1. Yapısı ve dağılımı
PRL’in aminoasit dizilimi, büyüme hormonu (GH) ve plasental laktojen (PL) ile benzerlik göstermektedir. Bu proteinler, benzer genomik, yapısal, immünolojik ve biyolojik özellikler gösterdiğinden, PRL/GH/PL protein ailesi olarak adlandırılmışlardır (28, 29). Yakın zamanda, PRL/GH/PL ailesinin, çok daha geniş bir aile olan hemotopoetik sitokin ailesi ile bağlantılı olduğu anlaşılmıştır (30). Prolaktinin reseptörü (PRLR) de, GH reseptörü, leptin reseptörü, eritropoetin (EPO) reseptörü ve pek çok interlökin (IL) reseptörünün mensubu olduğu, sınıf-1 sitokin reseptör ailesindendir (31). Bu reseptör, tek ve membran bağlı bir protein yapıdan oluşur (32). Ligandları gibi PRL ve GH’un reseptörleri de, benzer yapısal özellikler gösterir; 2
adet disülfid bağları, çift triptofan-serin dizilimleri mevcuttur (3). Her biri ekstraselüler, transmembran, intraselüler bölgeler içerir (3, 32). Anılan reseptörü kodlayan gen, 5.kromozom üzerinde yerleşmiştir, en azından 10 ekzon bölgesi vardır (31, 33). PRLR geninin transkripsiyonel düzenlenmesi -3 farklı, doku özgül olan- promoter bölge ile sağlanır. Promoter 1, gonadlar; promoter 2 karaciğer, promoter 3 ise gonadlar ve gonadların dışındaki organlara özgüdür (34). Farklı dokularda, PRLR geninin, uzunluğu ve kompozisyonu farklı olan izoformları mevcuttur (3, 4). Farklı izoformların ortaya çıkması, transkripsiyon ve alternatif ayrılma işlemlerinin, PRLR promoter bölgelerinin alternatif başlangıç bölgelerinde başlamasından kaynaklanır (35, 36). Östrojen yoğunluğuna, laktasyon durumuna ve gebeliğe bağlı farklı izoformların oluştuğu söylense de, PRLR’lerinin tüm vücuttaki yaygın dağılımı gözönünde bulundurulduğunda, farklı mekanizmaların da etkili olabileceğini düşünülmektedir (3). İzoformların sitoplazmik bölgelerinin uzunluğu ve kompozisyonu farklı olsa da, ekstraselüler bölgeleri benzerdir (3, 37). Membran bağlı reseptörlerin dışında, çözünebilir PRL-bağlayan proteinler de insan meme epitel hücrelerinde ve sütte gösterilmiştir (38, 39). PRLR’nün vücuttaki hemen tüm dokularda eksprese edildiği gösterilmiştir (3). Meme dokusu ve overler, PRLR’nün en iyi tanımlandığı bölgelerdir (40). Santral sinir sisteminde, pek çok bölgede; koroid pleksus, stria terminalis, amigdala, orta beyin, talamus, hipotalamus, serebral korteks ve olfaktör kanalda PRLR ve mRNA’sı gösterilmiştir. Periferik organlarda da, hipofiz, kalp, akciğer, timus, dalak, karaciğer, pankreas, böbrek, adrenal bez, uterus, iskelet kası, adipoz doku ve deride de PRLR mevcuttur (2-5, 40).
2.3.2. PRLR aktivasyonu
PRLR’nün, ekstraselüler bölgesinde ligand bağımlı dimerizasyon gerçekleşir (4). PRLR aktivasyonu, ligandın uyardığı ardışık reseptör dimerizasyonunun gerçekleşmesi ile başlar. Her bir PRL molekülü, 1. bölgenin 1 ve 4 numaralı heliksler, 2. bölgenin 1 ve 3 numaralı heliksler tarafından çevrelendiği 2 adet bağlanma bölgesi içerir. İlk olarak, 1. bağlanma bölgesi, PRLR ile etkileşime girer. Bu başlangıç ve hormon-reseptör kompleksinin oluşması, 2. bağlanma bölgesinin aynı PRL molekülü üzerinde diğer bir PRLR ile etkileşime girmesi için gereklidir (3).
Yirmidört aminoasit uzunluğundaki transmembran bölgesinin, PRLR aktivasyonundaki rolü bilinmemektedir (3, 4).
İntraselüler bölge ise, PRLR ile ilgili dönüşüm sinyal mekanizmalarının başlatılmasında anahtar rol oynar, diğer sitokin reseptörleri ile az miktarda dizilim benzerlikleri gösterir (3).
Bununla birlikte, PRLR üzerinde göreceli olarak korunmuş box 1 ve box 2 denilen iki alan bulunur (41). Box 1, reseptörün açılıp kapanması için gerekli olan dönüşüm sinyalleri tarafından tanınan, membranın proksimalinde bulunan, prolinden zengin bir yapıdır. Box 2, daha az korunmuştur, PRLR kısa izoformunda bulunmaz (3). PRLR’nün intraselüler bölgesi, herhangi bir intrensek enzimatik aktiviteden korunmuş olsa da, PRLR’nün ligandla düzenlenen aktivasyonu, reseptörün kendisi de dahil olmak üzere, pek çok selüler proteinin tirozinle fosforillenmesine neden olur (37, 42). İntraselüler bölgenin, membrana yakın bölgesi, Janus kinase 2 (Jak 2) denilen bir tirozin kinaz ile yakından ilgilidir (43-45). Jak 2 ve PRLR arasındaki ilişki, şüpheye yer bırakmayacak şekilde ispatlanmış olmakla birlikte, aralarındaki bağlantının yapısı tam olarak bilinmemektedir (4, 43, 45). PRL’in bağlanmasıyla, 1 dakika içinde, Jak 2’nin fosforilasyonunun gerçekleşmesi, Jak 2’nin, PRLR aktivasyonunda oldukça mühim bir rolü olduğunu düşündürmektedir (3, 43). Jak 2 aktivasyonu için, PRLR’nün intraselüler bölgesinde, prolinden zengin Box 1 ve ligandla uyarılmış PRLR dimerlerinin muhakkak bulunması gerekmektedir (43, 46-48). Jak 2 aktivasyonu, 2 adet Jak 2 molekülünü birbirine yakınlaştıran reseptör dimerizasyonu sonrası transfosforillenmeyle oluşur (48). Jak 2 kinazlar birbirlerini fosforiller iken, PRLR’nün tirozin rezidülerinin de fosforillenmesini sağlarlar (49). Fosfotirozinler, SH2 alanları içeren dönüştürücü moleküller için potansiyel bağlanma/havuzlanma alanları olmasından dolayı oldukça önemlidir (4, 50). İntraselüler bölgesinde 4 adet tirozin rezidüsü bulunmasına rağmen, reseptörün kendisinin tirozin fosforillenmesi PRLR’nün kısa formunun aktivasyonu sonrası görülmez (50). Proliferasyon gibi başlıca selüler fonksiyonlar, PRLR fosforilenmesi olmadan, PRLR kısa formuyla gerçekleşir (51). PRLR’nün uzun formu PRLR aktivasyonu sırasında fosforillenen pek çok tirozin rezidüleri içerir (52).
Jak 2 aktive olduğunda, farklı hedef proteinler üzerindeki tirozin rezidülerini fosforiller. Bunların en iyi bilinenleri, reseptörün bizzat kendisi ve Stat (Signal transducer and activator of transcription) adı verilen dönüştürücü proteinlerdir (3). Stat protein ailesi, sitokin reseptör sinyalizasyonunda, en önemli dönüştürücülerdir (44). Stat ailesinde 8 eleman bulunur; Stat 1, Stat 3, özellikle de Stat 5a ve Stat 5b, PRLR için başlıca dönüştürücü moleküller olarak kabul edilmektedir (53, 54). Stat molekülü, 5 adet korunmuş yapı içerir; DNA bağlayan bölge, SH3 benzeri bölge, SH2 benzeri bölge, NH2 ve karboksiterminal aktive edici bölgedir (55). İlk olarak, sitokin bağlı reseptör, ilgili Jak kinaz ile tirozin fosforilasyonuna uğrar. Ardından, fosforillenmiş tirozin, reseptör-Jak kompleksinin sonraki bölümünü oluşturmak üzere, bir Stat molekülünün SH2 bölgesi ile etkileşir. Üçüncü olarak, reseptöre bağlı Stat, komplekse ait olan
Jak kinaz tarafından fosforile edilir. Sonrasında, fosforillenmiş Stat, diğer Stat molekülünün SH2 bölgesi ve fosfotirozin içeren her monomerini kapsayan bir etkileşimle, homo veya heterodimerler oluşturarak reseptörden ayrılır. Son olarak, dimer, sitokin hedef genlerinin promoterlerinde bulunan özgün DNA elemanlarını aktive ettiği nükleusa geçer (3). Tirozin fosforilasyonu yanında, Stat aktivasyonu sonucu, serin/treonin fosforilasyonu da gerçekleşir. Stat 5a ve b izoformları arasındaki en önemli fark, serin/treonin fosforilasyon alanlarındaki farktır (56). Yeni elde edilen bilgiler, Stat 5’in, gen transkripsiyonunun düzenlenmesinde inhibitör rolünün olabileceğini göstermektedir (57).
Jak/Stat, en etkin yolaklar olmasına rağmen, mitojen aktive edici protein kinaz (MAPK) kaskadının da PRLR aktivasyonu için en az Jak/Stat kadar önemli olduğunu düşündüren çalışmalar mevcuttur (58-61). PRLR’nün fosfotirozin rezidüleri, reseptörü, Ras/Raf/MAPK kaskadına bağlayarak, adaptör proteinler (Shc/Grb2/SOS) için bağlayıcı alanlar olarak iş görebilirler (62, 63). Başlangıçta, Jak/Stat ve MAPK yolaklarının paralel veya bağımsız olduğu kabul edilirken, bu yolakların aslında birbiri ile bağlantılı olduğuna işaret eden birtakım bilgiler de mevcuttur (64).
Src kinaz ailesinin bir üyesi olan Fyn, PRLR ile ilişkilidir, PRL stimülasyonu ile aktive olduğu gösterilmiştir (65-67). Src kinazların PRLR’leri ile olan sinyal dönüşümündeki rolleri tam olarak bilinmese de, hücre büyümesini destekledikleri düşünülmüştür (65).
Yakın zamanda, insülin reseptör substratı 1 (IRS-1) ve fosfatidil inozitol (PI) 39-kinazın alt biriminin PRL ile uyarılan treonin fosforillenmesine uğradığı gösterilmiştir (68-70). Hem IRS-1, hem de PI-39 kinazın PRLR kompleksi ile ilişkili olduğu görülmektedir. PI-39 kinazın, PRL ile uyarılan aktivasyonunun Fyn ile düzenlendiği öne sürülmüştür (4).
PRLR’nün, en azından iki olayı ve iki bölgesi PRL ile uyarılan iyonik değişimlerle ilgili görünmektedir. PRLR’nün intraselüler bölgesindeki Box 1, tirozin kinaz bağımlı K1 kanallarının Jak 2 ile aktivasyonunda görev alır (71). İntraselüler bölgenin karboksi terminali, voltaj bağımsız Ca21 kanallarını açan hücre içi mesajcıların (inozitol 1,3,4,5-tetrakifosfat, inozitol hekzakifosfat) üretiminde rol alır (70, 72).
PRLR aktivasyonu, pek çok sinyal molekülünün tirozin fosforillenmesine neden olduğu için, sinyalizasyon yolaklarının inaktivasyonunun da tirozin fosfatazlarla ilgili olması beklenebilir (4).
Deneysel veriler, SH2 içeren tirozin fosfatazlar olan SHP-1 ve SHP-2’nin, PRL sinyalizasyonunda yavaşlatıcı rolleri olduğuna işaret etmektedir (73-75). Sitokin reseptör sinyalizasyonunun yeni ortaya konulan bir yönü, Jak/Stat yolaklarını inhibe eden SH2 içeren protein ailelerinin (CIS; cytokine-inducible SH2-containing family) ve sitokin sinyalizasyonunun baskılayıcılarının (SOCS; suppressors of cytokine signaling) keşfedilmesidir (76-80). Bu moleküllerin PRLR sinyalizasyonundaki asıl görevleri yakın zamanda tanımlanmıştır (76). PRL, SOCS-1 ve SOCS-3’ün akut ve geçici ekspresyonunu uyarmaktadır (75). SOCS-1 ve SOCS-3, Jak2’nin katalitik aktivitesini ve Stat proteinlerinin aktivasyonunu inhibe ederek, PRLR araclığıyla gerçekleşen sinyalizasyonu kapatırlar (75). CIS ve SOCS-2 genleri, PRL uygulaması ile ortaya çıkan uzamış aktivasyona cevap verirler, SOCS-2, muhtemelen, SOCS-1’in inhibitör etkisini baskılayarak, hücrelerin PRLR uyarısına duyarlılıklarını düzeltmektedir (75).
Fosfolipaz C (PLC; phospholipase C), protein kinaz C de PRLR sinyalizasyon yolağında rol aldığı düşünülen diğer enzimlerdir, fakat etkiledikleri substratlar net olarak ortaya konamamıştır (3, 81, 82).
2.4. PRL’in fonksiyonları
PRL’in ilk olarak meme gelişimini uyararak laktasyonu sağladığı keşfedildikten sonra, PRL ile ilgili olarak bulunan ikinci özellik korpus luteumun gelişmesindeki etkisi olmuştur (1, 83). Günümüzde, PRL’in, pek çok önemli biyolojik fonksiyonu daha olduğu ortaya çıkarılmıştır (2-5). Bir derlemede, PRL ile ilgili 300 farklı aktivite tarif edilmiştir (3). PRL’in etkilediği mekanizmaların başlıca şu alt başlıklar halinde toplanması mümkündür; 1-) su ve elektrolit dengesi 2-) büyüme ve gelişme 3-) endokrinoloji ve metabolizma 4-) beyin ve davranış 5-) üreme 6-) immünregülasyon ve koruma (2-5).
1-) Su ve elektrolit dengesi
PRL, neredeyse tüm omurgalılarda, su ve elektrolit dengesinde çok önemli bir role sahip iken, memelilerdeki fonksiyonları, diğer canlılardaki kadar net anlaşılamamıştır (3). Renal Na ve K atılımını azalttığı, adenozin trifosfataz’ı (ATPaz) artırdığı gösterilmiştir (84). PRL, terdeki Na ve Cl’u azaltmakta (85), ince barsağın tüm bölümlerinde, su ve tuz emilimini artırmaktadır (86). PRL, amnion sıvısını azaltıcı etki gösterir (87). Tüm bu veriler göz önünde bulundurulduğunda, PRL’in, gebeliğin geç dönemindeki artmış solüt transportunun, gebe anneyi laktasyon için hazırlamasına katkıda bulunduğu düşünülebilir (4).
2-) Büyüme ve gelişme
GH ve PRL’in kökleri ve yapısal özellikleri bakımından pek çok benzerlik göstermesi nedeniyle, PRL’in de büyüme üzerinde etkileri olabileceği düşünülmüştür, bu etkiler aşağı vertebralılarda, farklı dokularda gösterilmiş olmakla birlikte memelilerde, PRL’in büyüme üzerine direkt etkisi gösterilememiştir (3). Bazı çalışmalarda, PRL’in, hepatositlerin proliferasyonunda önemli bir role sahip olabileceği düşünülmüştür (3, 88). PRL’in intestinal mukoza hücrelerinin büyüklüğünü; vasküler düz kas hücresinin, pankreasın beta hücrelerinin, hipofizer GH3 hücrelerinin, insan benign prostat hipertrofisi epitel hücrelerinin, astrositlerin ve pek çok immün sistem hücresinin proliferasyonunu artırdığı gösterilmiştir (81, 82, 89-97). PRL’in, çeşitli gelişim basamaklarında da rolü olduğu düşünülmektedir. Akciğerlerin gelişimini, sürfaktan üretimini, preadiposit farklılaşmasını, germ hücrelerinin matürasyonunu, tüberoinfindübüler hipotalamik dopamin gelişimini artırdığı gözlenmiştir (98-102).
3-) Endokrinoloji ve metabolizma
PRLR’nün adipoz dokuda henüz saptanamadığı zamanlarda, PRL’in adipoz doku üzerinde indirekt etkili olabileceği düşünülürdü (103). Bugün PRLR’lerinin uzun ve kısa izoformları hem beyaz, hem de kahverengi yağ dokusunda gösterilmiştir. Uzun izoformu, Jak 2 sinyalizasyonunu ve Jak 2/Stat yolağının aktivasyonunu düzenlerken, kısa izoformu ya farklı yolakları kullanmakta, ya da dominant negatif inhibitör olarak fonksiyon görmektedir (104). PRL’in reseptörüne bağlanması, ardından dimerizasyonun ve Jak 2’nin fosforilasyonunun gerçekleşmesi ile Stat 5a/b çalışmaya başlar. Stat5a/5b nükleusa geçerek PRL’e cevap veren genlerin transkripsiyonunu değiştirir. Bunlar Jak 2/Stat’ın daha fazla aktive olmasını engelleyen SOCS ailesi üyeleridir. PRL, lipid metabolizmasını, adipoz dokuya serbest yağ asitleri (FFA; Free Faty Acids) sağlayan lipoprotein lipazı (LPL) ve malonil CoA’yı FFA’ya çeviren yağ asidi sentazı (FAS; Fatty Acid Synthase) inhibe ederek etkilemektedir. Malonil CoA, insülin duyarlı bir glukoz taşıyıcısı olan GLUT4 tarafından hücreye taşınan glukozdan elde edilir. İlave olarak, PRL, adiponektin ve IL-6 sekresyonunu da inhibe eder. Metabolik duruma bağlı olarak, leptini hem aktive, hem de inhibe edebilir. Resistin salınımı üzerinde bir etkisi yoktur (5). PRL’in adipogenez üzerinde etkileri olabileceğine dair çalışmalar vardır ancak bu konu henüz netlik kazanmamıştır (5).
PRL’in dopamin antagonistleri ile uyarılmış kronik yüksekliği ve günlük enjeksiyonu, artmış besin alımı ve vücut ağırlığı ile ilişkili bulunmakla birlikte, adipoz doku kitle artışı ile ilişkili
bulunamamıştır (105-108). PRL’in bromokriptin ile baskılanması, tersi bir etki yaratır (106, 109). Paraventriküler nükleusa PRL enjeksiyonu besin alımını artırmaktadır, bu da PRL’in periferik alanların yanında hipotalamik oroksejenik ve anoreksijenik bölgeleri de etkilediğini göstermektedir (5, 110).
PRL’in karbohidrat metabolizmasını farklı yollardan etkilediği gösterilmiştir, hiperglisemik, diyabetojenik etkileri olduğu bilinmektedir. Embden-Meyerhoff yolağı ve heksoz monofosfat şantı üzerinde etkileri vardır (111). İzole hepatositlerde, PRL’in fizyolojik konsantrasyonlarda, glikojen fosforilaz aktivitesini 4 kat artırdığı gözlenmiştir (112).
Erken fötal hayatta, PRLR’lerinin, islet asiner hücreleri ve kanallarında üretildiği gösterilmiştir, peri-postnatal dönemlerde insülin ve glukagon üreten hücrelerde daha baskın olduğu gözlenir (113). Gebelikte, pankreas islet hücrelerinde, PRLR ekspresyonu artar (114). Gebelikte glukozla uyarılan insülin sekresyonu eşiği düşer, beta hücrelerindeki 2 glukoz sensörü glukokinaz ve GLUT2, PRL ile uyarılır (115).
PRL, insülin gen transkripsiyonunu uyararak, insülin sekresyonunu uyarır. Hangi mekanizmalarla uyardığı tam olarak bilinmemekle birlikte, Stat5, PI3 kinaz, MAPK yolaklarını aktive ettiği gösterilmiştir (116, 117).
PRL’in, dihidroepiandrosteron, dihidroepiandrosteron sülfat, kortizol ve aldosteron gibi adrenal steroidlerin sentezini artırdığı gösterilmiştir (118, 119). PRL’in katekolaminlerin sentezini direkt olarak uyardığını gösteren çalışmalar olsa da, adrenal medullada PRLR gösterilemediğinden, bu etkinin artmış adrenokortikal steroid hormonlardan kaynaklanabileceği düşünülmektedir (3, 120, 121).
4-) Beyin ve davranış
Beyinde PRL’e atfedilen pek çok fonksiyon tanımlanmıştır. Bunlar; serebrospinal sıvının kompozisyonunun düzenlenmesi, astrositlerdeki mitojenik etkiler, serbestleştirici/baskılayıcı faktörlerin düzenlenmesi, uyku-uyanıklık sikluslarının düzenlenmesi ve üreme, ebeveyn olma, emzirme-besleme dürtülerinin düzenlenmesi olarak sayılabilir (2). Beyindeki PRL reseptörlerinin dağılımı ile PRL’in beyindeki fonksiyonları arasında net bir korelasyon kurulamamıştır (2).
Yeni doğum yapmış kadınların hemen hepsinde, bir çeşit annelik içgüdüsü, koruyucu, kollayıcı davranış paterni gözlenmektedir, hiç doğum yapmamış kadınlarda bu duygu daha az belirgindir, ya da hiç gözlenmez. Gebelik boyunca ve gebeliğin sonunda PRL ve PL’in artmış konsantrasyonlarının bu annelik güdülerini artırdığı düşünülmektedir (122-124).
PRL’in analjezik etki yapabildiği gösterilmiştir (125).
Kadınlarda, PRL’in yükselmesinin bazı psikomatik reaksiyonlara yol açabildiği gösterilmiştir, psödogebelik de bunların arasındadır (126).
Artmış PRL düzeylerinin, libidoda azalmaya, uyku-uyanıklık siklusunda değişikliklere neden olabileceği düşünülmektedir (126-128).
PRL’in, ventral hipotalamik nöronlardaki elektriksel aktiviteyi artırmak ve protein kinaz C aktivitesini güçlendirmek üzere, dopamin dönüşümünü artırdığı bilinmektedir (129, 130). Retinada, PRL’in iki etkisi gösterilmiştir; TRH reseptörlerinin sayısında azalmaya neden olur, fotoreseptör yıkımını artırır (131, 132).
5-) Üreme
PRL için tanımlanan fonksiyonlar içinde en geniş grubu, üreme ile ilgili olanlar kapsamaktadır (2). PRL’in en net tanımlanan etkileri memede gerçekleşenlerdir (2).
Meme dokusunun gelişiminde; hücresel büyüme, farklılaşma, sekresyon ve involusyonda, PRL, başrolü oynamaktadır. GH, PL, insülin, östrojen, progesteron, glukokortikoidler de gebelikte meme dokusunun gelişiminde rol oynayan diğer faktörlerdir. Meme dokusunun gelişiminde, son aşamada, tübüloalveolar gelişimi doğrudan PRL düzenlemektedir (2, 16). Emzirme dönemi boyunca, süt ihtiyacına göre, PRL, süt yapımı için gerekli faktörlerin, yağ dokudan memeye transferinde en önemli rolü oynar (133). Sütün içeriğindeki başlıca maddeler olan protein, laktoz ve lipidlerin sentezini düzenler (134). Memede yağların üretimini anahtar enzimleri koordine ederek sağlar. Dolaşan lipoprotein-trigliserid komplekslerinin hidrolizini ve yağ asitlerinin alımını sağlayan LPL’yi stimüle eder, asetil KoA’yı oluşturan piruvat dehidrojenazı aktive eder, malonil KoA üretiminde anahtar bir enzim olan asetil KoA karboksilazın ekspresyonunu artırır, yağ asidi sentezini artırır (135-137). PRL’in ayrıca, normal ve neoplastik meme dokusunda insülin benzeri madde 1 (IGF-1; Insulin like Growth Factor-1), epidermal büyüme faktörü (EGF; Epidermal Growth Factor),
glikolize musin, paratiroid benzeri peptid, PRL ile uyarılabilen proteinleri direkt uyarmaktadır (139-142).
Fertilize olmuş ovumun implantasyonu, gebeliğin sürdürülebilmesi, ovulasyonun inhibisyonu için overler tarafından üretilen progesteron gerekli olduğundan, progesteron üretiminin, korpus luteum tarafından düzenlenmesi oldukça önem taşır (143). PRL’in, luteotropik etkileri, luteal hücreler tarafından progesteron üretiminin uyarılmasını kapsar (144). Siklusun bulunduğu döneme bağlı olarak, PRL, korpus luteumun yıkımında da görev alabilir (145, 146). Granüloza hücrelerinde, PRL, östrojen sentezini ve P450 aromataz aktivitesini inhibe eder, Stat5 aktivasyonu üzerinden alfa 2 makroglobulini uyarır (147-149). PRL’in ovosit matürasyonu üzerine de etkili olabileceğini gösteren çalışmalar vardır (3).
Uterusta, PRL, progesteron reseptörlerinin seviyesini artırır (150, 151). Uterin sıvı kaybını uyarır, progesteron metabolizmasını yavaşlatır (152, 153). Blastosist implantasyonunu başlatır, lösin aminopeptidaz aktivitesini ve glukozamin sentaz aktivitesini artırır (154, 155). Prostoglandinlerin ve fosfolipaz A2’nin salınımını artırır (156).
Erkeklerde PRL’in fizyolojik rolü hakkındaki bilgiler net değildir (3).
6-) İmmünregülasyon ve koruma
PRL ile immün sistem arasında ilişki olabileceği, ilk olarak 1972 yılında, PRL olmayan farelerde, eksojen olarak verilen PRL’in timik fonksiyonları iyileştirmesi ile düşünülmüştür (157).
PRL’in, lenfositlerin, hormonal ve selüler immünitesini artırdığı, hipofizektomi sonrası gelişen anemi, lökopeni ve trombositopeniyi düzelttiği, antikor oluşumunu ve selüler proliferasyonu uyardığı gösterilmiştir (158-160). Selüler proliferasyonun, laboratuvar ortamındaki endotoksin bulaşıyla ilgili olabileceği söylense de, bu konu netlik kazanmamıştır (3). Proliferasyona olan etkisinin yanında, PRL’in, DNA sentezini ve c-myc ekspresyonunu artırabildiği gösterilmiştir (161). PRL, aynı zamanda, IL-2 ve EPO reseptör düzeylerini de artırmaktadır (97, 162, 163).
PRL’in, proliferasyonu uyarmasının yanında, lenfositlerin apopitozunu inhibe ettiği de gösterilmiştir (164). PRL uygulaması ile, artmış greft rejeksiyonu ilgili bulunmuştur (165).
Makrofajların aktivasyonunun GH ile ilgili olduğu düşünülürken, yakın zamanda, makrofaj aktivasyonu ve süperoksit anyon üretiminin PRLR ile de düzenlendiği ortaya konulmuştur (166). PRL’in, monoblastik büyümeyi azaltmada, IFN gamma ile birlikte sinerjistik etki gösterdiği, kanamadan sonra Kupffer hücrelerinde sitokin gen ekspresyonunu da artırdığı gözlenmiştir (167). PRL, timik-nurse hücre kompleksinde, polimorfonükleer hücrelerin doğrudan ve spontan migrasyonunu azaltır, lenfosit-epitelyal hücre adheziv etkileşimlerini düzenler, meme dokusunda Ig-A salgılayan plazma hücrelerini artırır (168-170). PRL, karaciğerde, faktör XII üretimini artırır (171).
Sistemik lupus eritematozus, romatoid artrit gibi, otoimmün hastalığı olan hastaların bir kısmında, hiperprolaktinemi gözlenmesi, PRL’in otoimmünite ile de ilişkili olabileceğini desteklemektedir (2-4, 16, 172).
Bleacha ve arkadaşları (174), 1980’li yıllarda, baskı, sıcak ve soğuk stresinin, dinitrofurobenzene karşı oluşan temas duyarlılığını artırmasında, PRL ve GH’un sorumlu olduğunu keşfetmişlerdir. Fizyolojik koşullarda, PRL, GH, IGF-1, fizyolojik veya çevresel stres faktörlerine karşı salgılanan ‘strese cevap veren, stres karşıtı hormonlar’ olarak bilinmektedir (172, 173). PRL salgılanmasını tetikleyen pek çok stres çeşidi tariflenmiştir; eter, baskı, termal, kanama, sosyal fobi, hatta akademik stres gibi (4).
7-) Diğer
PRL’in proteolitik fragmanlarının anjiogenezi inhibe edebildiği gösterilmiştir (175). Anjiogenez üzerindeki bu inhibisyonun fizyolojik önemi tam olarak bilinmese de, tümör oluşumunun engellenmesinde, terapötik amaçla kullanılabilecekleri düşünülmektedir (4).
PRL, bazı kanserlerle ilişkilendirilmiştir (3). PRL’in kolorektal kanser agresifliğini; meme kanserlerinde proliferasyonu; promiyelositlerin çoğalmasını artırdığı, malign B lenfositlerini ve lenfoma hücrelerini aktive ettiği gösterilmiştir (62, 176-182). Benign fibromüsküler tümörler olan leiomiyomların, normal miyometrium dokusundan daha fazla PRL ürettikleri gözlenmiştir (183).
2.5. PRL ve trombosit ilişkisi
2.5.1. Trombosit aktivasyonu, agregasyonu
Aktive olmuş trombositler, lökositlerle –özellikle monositlerle- bir araya gelerek birleşebilir ve agregatlar oluşturabilir (184). Bu agregatlar, koroner arter hastalığı olan kişilerin kanında yüksek oranlarda bulunmaktadır (185-187). Selektin ailesinin bir üyesi olan, P selektin –veya PADGEM, GMP-140, CD62P- 140 kDa ağırlığında bir glikoproteindir. Aktive olmayan trombositlerin yoğun granüllerinde ve endotelyal hücrelerin Weibel-Palade granüllerinde bulunur (188). P selektin, hücre adezyon kaskadında ilk basamak olan aktive endotel üzerindeki monositlerin hareketini düzenler. Trombositlerde, trombin, histamin veya C5a gibi agonistlerle aktivasyon sonrası, P selektin, granül membrandan, plazmatik membranın yüzeyine geçer ve trombositlerin nötrofil ve monositlere adezyonunu düzenler (189). P-selektin’in, lökositler üzerindeki bir ligandla (P selektin glikoprotein ligandı-1) etkileşimi, GPIIb/IIIa-fibrinojen (Glikoprotein IIb/IIa) etkileşimini stabilize ederek, büyük, kararlı trombosit agregatları oluşmasına yardım eder. Oluşan lökosit aktivasyonuyla, lökosit aktivasyonu için 2integrin Mac-1 denen başka bir yüzey reseptörünü aktive eder (190). Trombosit fonksiyonunda adenozindifosfat (ADP) çok önemlidir; diğer agonistler vasıtasıyla kolaylaştırılan cevabı ve P-selektin salınımını stimüle eder. ADP, depolanmış granüllerden salgılanan, zayıf fakat önemli agonisttir. Trombositlerdeki şekil değişikliğini ve geri-dönüşümlü trombosit agregasyonunu direkt olarak stimüle eder. Ardından ADP ile uyarılan tromboksan A2 sentezi oluşması ile trombositlerin ikincil agregasyonu başlar. Normalde hücre yüzeyinde eksprese edilmeyen P selektin, ADP ile stimüle edilince hücre yüzeyinde eksprese edilmeye başlanır (189).
Aktive trombositler, endotel aktivasyonu ve düz kas hücresi proliferasyonunu artıran proinflamatuvar sitokin ve düzenleyici büyüme faktörleri salgılarlar (187). Bu gözlemlerden yola çıkarak, aktive trombositlerden eksprese edilen P selektinin aterosklerotik lezyonların başlangıç aşamasında görev aldığı söylenmektedir (187).
Gebelik ve sonrası, venöz tromboemboliler (VTE) için riskli bir dönemdir. Gebelik sonrası erken dönemde, gebeliğe dair etkiler ortadan kalkmasına rağmen, VTE riski devam etmektedir. Bu nedenle, PRL ve VTE arasında, dolayısıyla da PRL ve trombosit aktivasyonu arasında bir ilişki olabileceği ileri sürülebilir. Bu tezden yola çıkılarak, gebelikteki VTE riskinin diğer risk faktörlerinin yanında, trombosit aktivasyonunun artmış olmasından kaynaklandığı düşünülmektedir (19, 191).
VTE insidansı yaklaşık, 1/1000’dir (192). VTE için bazı doğuştan gelen veya sonradan kazanılmış risk faktörleri olabilir. Bunların bazıları; anti-fosfolipid sendromu, hiperhomosisteinemi, aktif protein C ve S direnci, Faktör V Leiden mutasyonu, artmış faktör VIII düzeyidir. VTE gelişimini kolaylaştırdığı düşünülen diğer bazı durumlar gebelik, tümörler, ileri yaş, cinsiyet, cerrahi ve travmadır (192).
VTE, gebelik boyunca ve lohusalık döneminde önemli bir mortalite ve morbidite nedeni olarak önemini korumaktadır (193). Olguların yaklaşık %50’sinde, hareketsizlik veya doğuştan bazı risk faktörleri ile VTE’den sorumlu olabilir, geri kalan -idyopatik- hastalarda neden açıklanamamaktadır (192, 193).
Bu durum, araştırmacıları, gebelik ve doğum sonrasında VTE’ye neden olan diğer faktörleri araştırmaya itmiştir. Gebelik boyunca artan ve gebelik sonrasında da yüksek kalan PRL düzeylerinin, gebelik boyunca trombosit aktivasyonunda bir kofaktör olarak görev alabileceği düşünülmüştür. Flovsitometri tekniklerinin ilerlemesi ile trombosit agregasyonunda rol alan P- selektin salınımı trombosit yüzeyinde gösterilmiştir (191).
PRL ile trombositlerin ilişkisi hakkında çok fazla veri yoktur. Hipofizektomi yapılan ratlarda, kemik iliğinde görülen trombositopeninin dışarıdan PRL verilmesi ile düzeltilebildiği gösterilmiştir (160). Dardenne ve arkadaşları (194), 1994 yılında yaptıkları çalışmada, PRL ve PRLR’nü hemopoetik dokularda göstermişlerdir, bununla birlikte PRL ve PRLR’nün hemopoetik hücrelerin üzerindeki etkileri netlik kazanmamıştır (195). Yine antipsikotik kullanan hastalarda, artmış VTE riskinin, antipsikotik kullanımına bağlı PRL yüksekliği ile ilişkili olabileceği ortaya atılmıştır (191, 196, 197). Tüm bunlardan yola çıkarak, 2001 yılından itibaren bazı çalışmalar, literatürde yerini bulmuştur (191, 197-201).
Wallaschofski ve arkadaşlarının (191), yaptığı çalışmada, trombositlerin ADP ile stimülasyonunun, PRL düzeyleri ile korelasyon gösterdiği; hiperprolaktineminin, trombosit agregasyonu için bir risk faktörü olabileceği; gebelikte artmış VTE riskine PRL yüksekliğine bağlı oluşan trombosit aktivasyonunun katkıda bulunabileceği öne sürülmüştür. Aynı grup, yaptıkları çalışmada, 1 yıl sonra, insan trombositleri üzerinde, monoklonal antikorlar kullanarak, PRLR ailesinin 43 kDa’luk kısa izoformunu göstermeyi başarmışlardır (198). ADP ile uyarılan trombosit agregasyonu, Gq ve Gi proteini ile düzenlendiğinden, PRL ile
ilişkisine bakılmış, PRL’in etkisini, Gq proteini aracılığıyla gösterdiği gözlenmiştir. ADP ve PRL’in, Gq aracılı sinyalizasyon yolağında anahtar bir enzim olan protein kinaz C’yi sinerji içinde aktive ettiği gösterilmiştir (198). Trombosit aktivasyonunda önemli rolü olan bazı yolakların –Ras/Raf/MAPK, PI-3 kinaz- PRL reseptörleri ile de ilişkisi olduğundan, PRLR aktivasyonu ile trombosit aktivasyonu arasında da ilişki olabileceği öne sürülmüştür (198). Yakın dönemlerde, antipsikotik kullanan hiperprolaktinemik hastalarda trombosit aktivasyonunun, normoprolaktinemik hastalara göre arttığı gösterilmiştir (197). İskemik inme geçiren 36 hastanın, serum PRL düzeyleri ile ADP ile uyarılmış trombosit aktivasyonları arasında pozitif bir korelasyon bulunmuştur (201). Ne var ki, PRL ile trombosit aktivasyonu arasındaki ilişki ile ilgili bilgiler henüz oldukça yenidir ve bu konuda daha fazla çalışma yapılmasına ihtiyaç vardır (191, 197-201).
2.6. Hiperprolaktinemi 2.6.1. Hipofiz kaynaklı
Hiperprolaktinemi, hipofizer hastalıklar içinde en sık görülenidir. Klinik semptomlara yol açan hiperprolaktinemi, daha çok, PRL salgılayan hipofizer bir adenoma bağlı ortaya çıkar (16, 202). Prolaktinomalar, tüm hipofizer tümörlerin %40’ını oluşturan iyi huylu tümörlerdir. Hastaların %30-80’inde galaktore vardır (202). Galaktore ve amenore kadınlarda en sık görülen semptomlardır, PRL’in meme ve hipotalamus-hipofiz-over aksındaki direkt etkisine bağlıdır (16). Kadınlarda, azalmış libido, vajinal kuruluk, disparoni diğer görülebilecek semptomlar iken, erkeklerde, impotans, vücut kıllarında azalma, testislerde yumuşama, infertilite ve jinekomasti görülebilir (202). Kadınlarda genellikle mikroadenomalar görülürken, erkeklerin büyük kısmı, makroadenomlar ve buna bağlı bası semptomları ile başvururlar (16, 202).
Hipofiz sapında baskı ya da kesilmeye neden olan herhangi bir kafa travması veya kafa içi bir kitle de hiptalamustan gelen dopaminerjik tonüsün kaybolmasına neden olacağından, hiperprolaktinemiye neden olabilir (16). Yine aynı şekilde, dopamin sentezini engelleyen herhangi bir neden de PRL değerinde yükselmeye neden olabilir (202).
2.6.2. Hipofiz dışı nedenler
Hafif hiperprolaktinemi, polikistik over sendromu (PKOS) olan kadınların %30’unda mevcuttur (203). PKOS ile hiperprolaktinemi antiteleri arasında belirgin bir etki veya
benzerlik veya ilişki bulunamamıştır (204). Bununla birlikte, PKOS’lu hastaların bir kısmında, hiperprolaktinemi varlığından bağımsız olarak, dopamin agonistleri verilmesi ile, bir grup olguda amenorenin düzeldiği gösterilmiştir (205). Meme başının uyarılması da kan PRL düzeylerini hafif de olsa yükseltebilir (16). Hipotiroidisi olan hastaların yaklaşık %20’sinde PRL düzeyleri yükselebilir. Sebep tam olarak bilinmese de, artmış TRH düzeylerinin PRL yüksekliğine neden olduğu düşünülür. Hipotiroidinin tedavisi ile PRL düzeyleri de normale dönebilmektedir (206). Kronik böbrek yetmezliği olan hastalarda, glomerüler filtrasyon hızınını azalmasına bağlı hiperprolaktinemi görülebilir (207). Emosyonel stres de daha önce bahsedildiği gibi, PRL düzeylerinde yüksekliğe neden olabilir (4,16). Bazı ilaçlar, hafif veya ılımlı PRL yüksekliğine neden olarak, galaktore, amenore, seksüel fonksiyon bozukluğu gibi hiperprolaktinemi semptomlarına neden olabilir (4, 202).
2.6.3. Makroprolaktinemi
Rogol ve Rosen (208), Suh ve Frantz (209), 1974 yılında, hiperprolaktinemik bazı hastalarda, big PRL’in baskın olan form olabileceğini ortaya koymuşlardır. Whittaker ve arkadaşları (210), 1981 yılında, hiperprolaktinemi varlığında fertilitenin korunabildiği hastalarda, ölçülen PRL’in büyük kısmını big big PRL’in oluşturduğunu göstermişlerdir. Hattori ve arkadaşları ile, Leite ve arkadaşlarının, aynı zamanlarda, idyopatik hiperprolaktinemisi olan hastaların bazılarının serumunda, anti-PRL antikorlarını göstermesi ile, makroprolaktin molekülünün, Ig ile kompleks yapmış monomerik PRL’den oluşan bir makromolekül kompleksi olduğu anlaşılmıştır. Molekül ağırlığı 23 kDa olan bir PRL ve Ig kompleksidir (11-13). Makroprolaktin kompleksinin PRL reseptörüne bağlanma afinitesi ve serumdan temizlenme hızının, monomerik PRL’e göre daha düşük olduğu gösterilmiştir. Makroprolaktinemisi olan kişilerde, makroprolaktinin birikerek kanda PRL değerinin yüksek ölçülmesine neden olduğu düşünülür (12, 211). Şimdiye kadar yapılan çalışmalar, makroprolaktinin in vivo biyoyararlanımı ve biyolojik etkinliğinin oldukça düşük olduğunu işaret etmektedir (212). Bu nedenle PRL yüksek olmasına rağmen, PRL yüksekliği ile ilgili klinik bulguların (amenore, galaktore, inferilite gibi) olmaması durumunda, makroprolaktinemi akla gelmelidir (212).
Makroprolaktineminin sıklığı tam olarak bilinmemektedir. Hiçbir yakınması olmayan, 10550 sağlıklı kişide yapılan tarama sonucunda, 40 hastada hiperprolaktinemi, hiperprolaktinemik hastaların ise 10’unda makroprolaktin pozitifliği tespit edilmiştir (213). İskandinavya’dan yapılan bir çalışmada, Bjoro ve arkadaşları (214), 660 sağlıklı birey içinde sadece bir kişide makroprolaktin tespit etmişlerdir. Çalışmalar gözönünde bulundurulduğunda,
makroprolaktinemi prevalansının genel toplumda; kadınlarda %0,2, erkeklerde %0,02 olabileceği tahmin edilmektedir. Hiperprolaktinemik popülasyonda, ölçümde kullanılan yönteme göre değişmekle birlikte, makroprolaktinemi sıklığı farklı çalışmalarda %10 ile %46 arasında değişebilmektedir ve bu farkın çalışma popülasyonunun seçiminden kaynaklandığı düşünülmektedir (212, 215, 216). Makroprolaktin ölçümü için çeşitli yöntemler kullanılmaktadır. Jel filtrasyon kromotografisi (JFK) bugün halen altın standart olarak kabul edilmektedir (212, 217). Çalışma esası moleküler ağırlığına göre proteinlerin ayrıştırılmasına dayanmaktadır. Makroprolaktin oranı, tüm PRL’in %40’unun üzerinde olduğunda, örnek makroprolaktin için pozitif olarak kabul edilmektedir (217). Bu yöntem güvenilirliği çok yüksek olmakla birlikte, bazı dezavantajlara da sahiptir (212). İlk olarak, Hattori ve arkadaşlarının da tarif ettiği gibi (12), düşük afiniteye sahip olan PRL antikor kompleksinde, oldukça uzun süren JFK işlemi sırasında, antikor ile PRL molekülünün birbirinden ayrılma, dolayısıyla da makroprolaktin miktarının olduğundan daha az ölçülme ihtimali mevcuttur. İkincisi, varolan makroprolaktin yüzdesi ile ilgili ölçümler konusunda belirsizlikler vardır. Diğer bir neden, JFK sırasında, adsorbsiyon ve denatürasyon işlemleriyle birlikte PRL immünreaktifinin işleme bağlı olarak kaybı, her bir farklı izoformun yanlış bir şekilde fazla ya da eksik olarak ölçülmesine neden olabilir. Son olarak JFK’nın, oldukça pahalı ve zaman alıcı bir yöntem olması nedeniyle yaygın olarak kullanımı kısıtlıdır (12).
Hattori ve arkadaşlarının (11), 1992 yılındaki tariflerinden beri, polietilen glikol (PEG) ile çöktürme yöntemi de oldukça yaygın olarak kullanılan bir testtir. Serum örneğinin PEG ile karıştırılıp ardından santrifüj edilmesinden sonra, çökme olmayan serumda ve işlem görmemiş serumda PRL ölçülür. Aradaki fark makroprolaktin düzeyini verir; %40’ın üzerinde çökme olması makroprolaktin varlığına işaret eder (217). PEG yöntemi özgün bir yöntem olmamasına rağmen JFK’ya göre oldukça hızlı ve ucuz (27 kat) bir yöntemdir, uygulaması kolaydır (12, 217). JFK ve PEG sonuçları korelasyon göstermektedir ancak kantitatif makroprolaktin düzeyleri arasında fark olabileceği gösterilmiştir (12). Yakın zamanda, Prazeres ve arkadaşları (218) ultrafiltrasyon metodunu keşfetmişlerdir.
2.7. Metabolik sendrom, insülin direnci, trombosit aktivasyonu 2.7.1. Metabolik sendrom ve insülin direnci
İnsülin direnci, fizyolojik konsantrasyonlarda üretilen insüline normal biyolojik yanıtın bozulması durumudur. İnsülinin biyolojik etkisini gösterebilmesi için, pankreas beta hücrelerinde sentez edilip salınması, portal sistem yoluyla sistemik dolaşıma katılması,
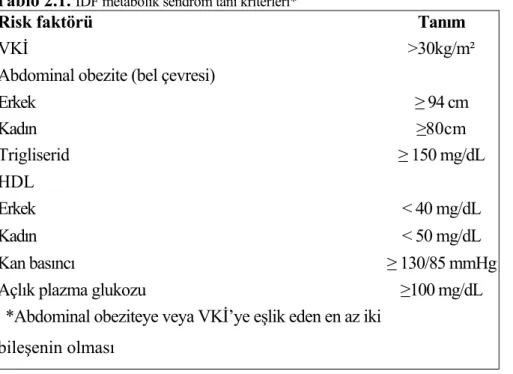
dolaşımdan interstisyuma geçmesi ve hedef dokulara ulaşarak bu doku hücrelerinin membranlarında bulunan spesifik reseptörlerle ilişkiye girmesi gerekmektedir. Reseptörü ile birleşen insülin, hormon etkisini gerçekleştirecek bir seri postreseptör olayı tetikleyecektir. Bu basamakların herhangi birinde veya birkaçında gerçekleşebilecek bir aksama, sonuçta organizmanın insüline subnormal yanıt vermesiyle sonuçlanacaktır (219). Metabolik sendromun, insülin direnci temelinde gelişen bir antite olduğu düşünülmektedir. İlk olarak, 1988 yılında, Reaven (220), insülinin in vivo etkisinin bozulması ile başlayan metabolik anomalilerin, dislipidemi ve hipertansiyon ile birlikte kardiyovasküler olaylara yol açtığını göstermiş ve bu antiteyi “sendrom x” olarak tanımlamıştır. Trigliserid yüksekliği, HDL-kolesterol düşüklüğü, hipertansiyon ve insülin direnci ile birlikte artmış insülin değerleri, sendromun bileşenleri olarak sayılmıştır. Uluslararası Diyabet Federasyonu (IDF), 2005 yılı başlarında metabolik sendrom tanı kılavuzunda, temel olarak abdominal obeziteyi kriter almış ve ona eşlik edecek dört komponentten ikisinin müspet olmasını tanı için gerekli bulmuştur. Vücut kütle indeksinin (VKİ), 30kg/m² olması durumunda ise, abdominal obeziteye bakılmasına gerek olmadığı bildirilmiştir. (Tablo 1) (221).
Tablo 2.1. IDF metabolik sendrom tanı kriterleri*
Risk faktörü Tanım
VKİ >30kg/m²
Abdominal obezite (bel çevresi)
Erkek ≥ 94 cm Kadın ≥80cm Trigliserid ≥ 150 mg/dL HDL Erkek < 40 mg/dL Kadın < 50 mg/dL Kan basıncı ≥ 130/85 mmHg
Açlık plazma glukozu ≥100 mg/dL
*Abdominal obeziteye veya VKİ’ye eşlik eden en az iki bileşenin olması
2.7.2.Metabolik sendrom, insülin direnci ve PRL
PRL’in bilinen etkilerinin yanında karbonhidrat ve yağ metabolizması üzerine olan etkileri keşfedildikten sonra, bu konuda pek çok çalışmalar yapılmıştır. PRL’in insülin direncine neden olduğu, diyabetojenik bir hormon olarak kabul edilebileceği, daha önce de bahsedildiği
gibi uzun zamandır bilinmektedir (5). Hiperprolaktinemi, adipoz doku artışı ile doğrudan ilgili bulunmasa da (105-108), hiperprolaktinemik hastaların, tedavi ile kilo kaybettikleri ve endotel fonksiyonları, metabolik parametreleri ve insülin dirençlerinin düzeldiği gösterilmiştir (222-225).
2.7.3.Metabolik sendrom, insülin direnci, trombosit aktivasyonu
Kan P selektin düzeyleri ile metabolik sendrom ve insülin direnci arasındaki ilişkiyi inceleyen son dönemde yapılmış pek çok çalışma mevcuttur. Obez hastalarda, kontrol grubuna göre yüksek olan P selektin düzeylerinin, diğer inflamatuvar belirteçlerle birlikte, kilo kaybı sonrası düştüğünü gösteren çalışmalar mevcuttur (226, 227). Dislipidemik bir popülasyonda yapılan çalışmada, kontrol grubuna göre kan P selektin düzeyleri, anlamlı olarak yüksek bulunmuşken, aynı popülasyonda hastaların insülin direnci değerlendirilmiş ve insülin direnci ile P selektin düzeyleri arasında bir ilişki bulunamamıştır (228). Metabolik sendromla kan P selektin ilişkisinin irdelendiği bir çalışmada, metabolik sendromu olan hastalarda, P selektin yüksek bulunmuştur (229). Başka bir çalışmada, farklı etnik gruplar incelenmiş, tüm popülasyonda metabolik sendrom kriterleri değerlendirilmiş, sistolik kan basıncı, HDL kolesterol ve trigliserid parametreleri ile P selektin düzeyleri arasında anlamlı ilişki bulunmuştur (230). Pergola ve arkadaşlarının (231), yaptığı çalışmada, hafif kilolu ve obez kadınlarda P selektin düzeyleri anlamlı olarak yüksek bulunmuş, P selektin düzeylerinin, VKİ, insülin direnci, sistolik ve diyastolik kan basıncı, açlık insülini, trigliserid ve PAI-1 düzeyleri ile pozitif korelasyon gösterdiği bulunmuştur. Ancak, çoklu regresyon analizi sonucunda, metabolik sendrom kriterlerinin, P selektin için bir risk faktörü olmadığı ortaya konmuştur. Çeşitli çalışmalarda, koroner arter hastalığı ve atriyal fibrilasyon için P selektinin bir risk faktörü olarak kabul edilmesi gerektiği yönünde farklı düşünceler ileri sürülmüştür (187, 230, 232).
3. HASTALAR VE YÖNTEM
3.1. Çalışma GrubuBu çalışma, Başkent Üniversitesi Tıp Fakültesi, Endokrinoloji ve Metabolizma Hastalıkları Bilim Dalı Kliniği’nde 2006-2007 tarihleri arasında yapıldı. Çalışmaya, PRL yüksekliği tespit edilen, daha önce hiç tedavi almamış 32 kadın hasta alındı. Hipofizde adenomu olan hiperprolaktinemik hastalarda diğer hipotalamo-hipofizer aksların normal çalıştığı gösterildi. Kontrol grubu olarak hastanemize genel kontrol amacıyla başvuran ve yapılan tetkiklerde herhangi bir sorun tespit edilmeyen, serum PRL düzeyleri normal sınırlarda, yaşı ve vücut kitle indeksleri hasta grubundan farklılık göstermeyen 33 sağlıklı gönüllü kadın birey dahil edildi.
Çalışmaya alınan hasta grubunda ve kontrol grubundaki bireylerin trombosit fonksiyonlarını etkileyecek herhangi bir ilaç kullanmamaları, sigara kullanmamaları, alkol kullanmamaları şartı arandı. Olguların anamnezlerinde ve aile öykülerinde trombotik bir olay olmamasına özen gösterildi.
KA 05/85 proje numarası ile Başkent Üniversitesi Tıp Fakültesi Etik Kurulu onayı alındı. Bireylere gönüllü denek bilgilendirme formu okutularak onayları alındı.
3.2. Yöntem
Hastaların başvuru nedenleri, boyları, kiloları, yaşları, bel çevreleri kaydedildi.
Serum PRL düzeyleri kemiluminesan mikropartikül immunoassay CMIA (Architect Ci8200, Abbott Laboratories, Diagnostics Division, Abbott Park, Illinois 60064, Referans Aralık: 1.20-29.93 ng/mL (SI units), 25.2-628.53mIU/L) yöntemi ile ölçüldü. Makroprolaktin ölçümü için hasta grubundan düz biyokimya tüpüne alınan kanlar, pıhtılaştıktan sonra santrifüj edilerek serumları ayrıldı ve -20◦C’de yaklaşık 10 ay saklandı. Örneklerden prolaktin tayini yapıldı. Makroprolaktin konsantrasyonunu tayin etmek için, örnekler polietilen glikol (PEG 8000-BioBasic Inc 3185 B10) ile ön işlemden geçtikten sonra tekrar prolaktin tayini yapıldı. Ikiyüzelli µl serum, 250 g/L pH 7.4’de fosfat tamponu (137 mmol/L sodyum klorid, 10 mmol/L sodyum fosfat) içinde çözünmüş PEG ile karıştırıldı, 10 dakika oda ısısında inkübe edildi. Karışım 3500 rpm’de 30 dakika santrifüj edildi, süpernatandan prolaktin tayini yapıldı. PEG ile ön işlem yapılmadan önce ölçülen prolaktin ile, PEG ile çöktürme sonrası ölçülen
prolaktin değeri arasındaki fark, makroprolaktin düzeyi olarak kaydedildi. Normal bireylerde PEG ile işlem sonrası %40’ın altında bir çökme beklenmektedir.
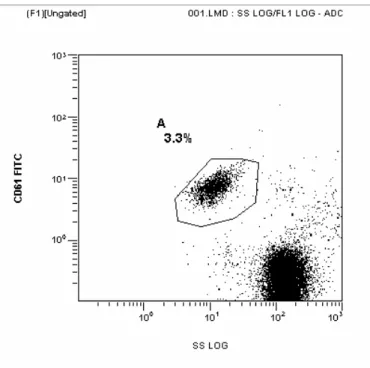
Stimüle edilmemiş ve ADP ile stimüle edilmiş P selektin düzeylerine bakıldı. P selektin, normal trombositlerde eksprese edilmediğinden ADP ile stimülasyon yaparak P selektin düzeyleri değerlendirildi. Hiperprolaktinemisi olan hastaların ve kontrol grubunun sitratlı tüpe alınan tam kanları % 0,5 bovin serum albuminli (Sigma) fosfat tamponu ile trombosit sayısı 20.000/μl olacak şekilde dilüe edildi. Dilüe edilen trombositlerle 2 ayrı çalışma yapıldı. Birinci çalışmada ADP (Sigma) sitimülasyonu yapılmaksızın akım sitometride direkt trombosit aktivasyonuna bakıldı. Bu amaçla tüpe 20 μl dilüe tam kan, 10μl anti CD61- FITC (Beckman Coulter, Marseille, France) ve 10μl anti CD62P- PE (Beckman Coulter, Marseille, France) eklendikten sonra 20 dakika oda sıcaklığında ve karanlıkta inkübe edildi. Diğer çalışmada ise; tüpe, hazırlanan trombosit süspansiyonundan 20μl ve 10μM ADP (final konsantrasyon) eklendi ve 37 0C’de 5dakika inkübe edildi. Süre sonunda 10μl anti CD61-FITC ve 10μl anti CD62P- PE monoklonal antikorları eklendi ve 20 dakika oda sıcaklığında ve karanlıkta inkübe edildi. İnkübasyon sonunda tüpler hiç bekletilmeden akım sitometri cihazında (Beckman Coulter EPICS XL-MCL) okutuldu. Sonuçlar, EXPO 32 ADC software kullanılarak analiz edildi. Trombositler anti CD61 kullanılarak seçildi ve non-spesifik bağlanmalar ekarte edildikten sonra anti CD62P ile aktivasyon oranları yüzde olarak tespit edildi. Trombositlerin seçilme işlemi Şekil 3.1’de gösterilmiştir.
P selektinin metabolik sendrom için bir risk faktörü olabileceği yönündeki farklı görüşler dikkate alınarak, hasta ve kontrol gruplarının metabolik sendrom parametreleri de çalışıldı. Olguların kan basınçları, standart cıvalı manometre ile, 15 dakikalık istirahati takiben yatar pozisyonda, sağ koldan, muayene başlangıcında ve bitiminde ölçüldü, ortalamaları alındı. Bel çevreleri IDF’e göre, mezura ile, olgular ayakta ve çıplak dururken, en alt kosta ile iliyak kenarın üst kenarı arasındaki bölgenin ortası ölçülerek elde edildi.
Açlık glukozları enzimatik kolorimetrik tayinle glukoz oksidaz yöntemi ile bakıldı. HDL kolesterolleri ve LDL kolesterolleri homojen enzimatik kolorimetrik yöntemle ölçüldü. Trigliseritleri, enzimatik kolorimetrik tayin yöntemi ile ölçüldü. Glukoz, HDL, LDL, trigliserit değerleri Roche MODULAR DP biyokimya analizörü cihazında çalışıldı. Açlık insülin düzeyleri, mikropartikül enzim immünoasay yöntemi ile, Axsym (Abbott Diagnostic Division) cihazı ile çalışıldı. Tüm olgulara, 10-14 saat açlık sonrası, sabah açlık kanının alınmasını takiben 75 g glukoz (300 ml suda çözünmüş) 5 dakika içinde içirilerek oral glukoz tolerans testi yapıldı. 120. dakikada kan örnekleri alınarak glukoz düzeyleri çalışıldı. Insülin dirençleri, HOMA-IR (Homeostasis Model Assesment) formülü ile hesaplandı; {Açlık Glukoz (mmol/l) X Açlık İnsülin (μU/ml) } / 22.5.
3.3. İstatistik
İstatistiksel değerlendirme için Windows SPSS 12.0 paket programı kullanıldı. p<0,05 istatistiksel olarak anlamlı olarak kabul edildi. Kullanılan istatistiksel testler: Deskriptif analiz, Bağımsız örnekler T-testi, Ki-kare ve Pearson korelasyon analizi idi.
4. BULGULAR
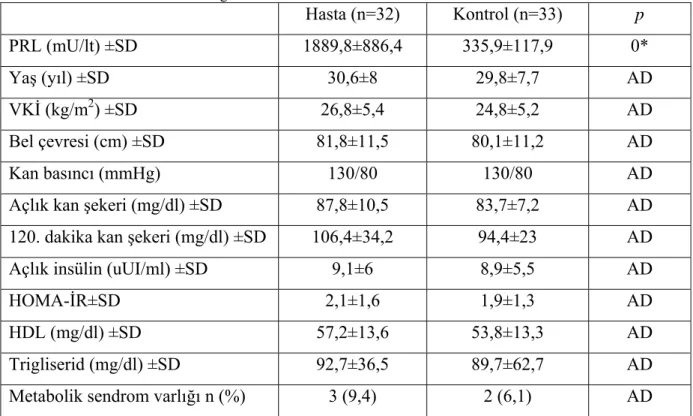
Çalışmaya 32 hasta, 33 kontrol grubu alındı.Hastaların ve kontrol grubunun karakteristik özellikleri ve laboratuvar verileri Tablo 4.1.’de verilmiştir.
Tablo 4.1. Hastaların ve kontrol grubunun karakteristik özellikleri ve laboratuvar verileri
Hasta (n=32) Kontrol (n=33) p PRL (mU/lt) ±SD 1889,8±886,4 335,9±117,9 0* Yaş (yıl) ±SD 30,6±8 29,8±7,7 AD VKİ (kg/m2) ±SD 26,8±5,4 24,8±5,2 AD Bel çevresi (cm) ±SD 81,8±11,5 80,1±11,2 AD Kan basıncı (mmHg) 130/80 130/80 AD
Açlık kan şekeri (mg/dl) ±SD 87,8±10,5 83,7±7,2 AD 120. dakika kan şekeri (mg/dl) ±SD 106,4±34,2 94,4±23 AD
Açlık insülin (uUI/ml) ±SD 9,1±6 8,9±5,5 AD
HOMA-İR±SD 2,1±1,6 1,9±1,3 AD
HDL (mg/dl) ±SD 57,2±13,6 53,8±13,3 AD
Trigliserid (mg/dl) ±SD 92,7±36,5 89,7±62,7 AD
Metabolik sendrom varlığı n (%) 3 (9,4) 2 (6,1) AD AD: Anlamlı değil
IDF kriterleri göz önünde bulundurularak, hastaların metabolik sendrom durumları değerlendirildi. Çalışma grubunda 3, kontrol grubunda 2 kişide metabolik sendrom mevcuttu, gruplar arasında metabolik sendrom mevcudiyeti açısından anlamlı fark yoktu. Diyabet açısından aile öyküleri sorgulandı, gruplar arasında anlamlı fark yoktu. PRL değerleri ile metabolik sendrom kriterleri arasındaki korelasyonda anlamlı ilişki izlenmedi.
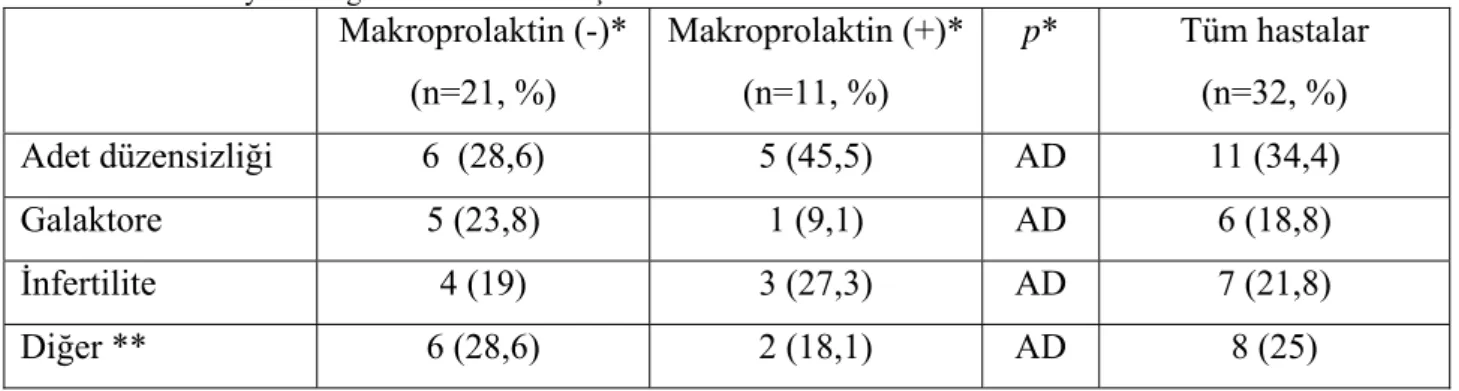
Hiperprolaktinemisi olan tüm hastalarda makroprolaktin ölçümü yapıldı, %40’ın üzerinde çökme olanlar makroprolaktin pozitif olarak kabul edildi, hastaların 11’inde (%29) makroprolaktin pozitif olarak bulundu. Makroprolaktin (+) grupla, makroprolaktin (-); gerçek hiperprolaktinemik grup arasında bazal PRL değerleri açısından fark bulunamadı (Tablo 4.2.). Çalışma grubunun makroprolaktin pozitifliği olan ve gerçek PRL yüksekliği olan alt-grupları
arasında da metabolik sendrom kriterleri ve metabolik sendrom varlığı açısından fark bulunamadı.
PRL yüksekliği olan hastalarda, kliniğe başvuru nedenleri ve hastaların hipofiz manyetik rezonans (MRG) görüntüleme sonuçları değerlendirildi (Tablo 4.2., Tablo 4.3).
Tablo 4.2. PRL yüksekliği olan hastalarda başvuru nedenleri
Makroprolaktin (-)* (n=21, %) Makroprolaktin (+)* (n=11, %) p* Tüm hastalar (n=32, %) Adet düzensizliği 6 (28,6) 5 (45,5) AD 11 (34,4) Galaktore 5 (23,8) 1 (9,1) AD 6 (18,8) İnfertilite 4 (19) 3 (27,3) AD 7 (21,8) Diğer ** 6 (28,6) 2 (18,1) AD 8 (25)
*İki grup karşılaştırılmıştır
**Diğer: saç dökülmesi, tüylenme, tesadüfen saptananlar, hipofiz insidentalomu
Tablo 4.3. PRL yüksekliği olan hastalarda (MRG) bulguları
Makroprolaktin (-) (n=21, %) Makroprolaktin (+) (n=11, %) P Mikroadenom 11 (52,5) 5 (45,5) AD Makroadenom 4 (19) 0 AD
Parsiyel empty sella (PES) 2 (9,5) 0 AD
Normal 4 (19) 6 (54,5) 0,04*
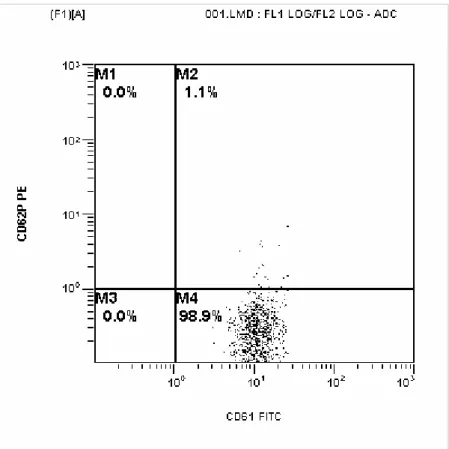
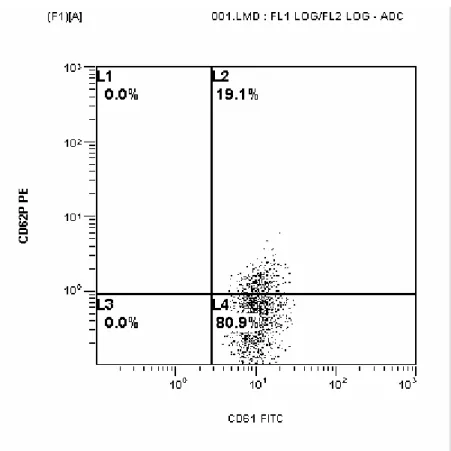
Kontrol grubu, gerçek hiperprolaktinemi grubu ve makroprolaktinemik grubun P selektin ekspresyonlarının örnekleri Şekil 4.1., 4.2. ve 4.3.’de gösterilmiştir.
Şekil 4.1. Kontrol grubunda P selektin ekspresyonu